
The Project Gutenberg EBook of Texas Fossils: An Amateur Collector's
Handbook, by William H. Matthews III
This eBook is for the use of anyone anywhere in the United States and most
other parts of the world at no cost and with almost no restrictions
whatsoever. You may copy it, give it away or re-use it under the terms of
the Project Gutenberg License included with this eBook or online at
www.gutenberg.org. If you are not located in the United States, you'll have
to check the laws of the country where you are located before using this ebook.
Title: Texas Fossils: An Amateur Collector's Handbook
Texas Bureau of Economic Geology Guidebook 2
Author: William H. Matthews III
Release Date: January 5, 2018 [EBook #56315]
Language: English
Character set encoding: UTF-8
*** START OF THIS PROJECT GUTENBERG EBOOK TEXAS FOSSILS ***
Produced by Stephen Hutcheson, MFR and the Online
Distributed Proofreading Team at http://www.pgdp.net
BUREAU OF ECONOMIC GEOLOGY
The University of Texas at Austin
Austin, Texas 78712
John T. Lonsdale, Director
Guidebook 2
By
William H. Matthews III

November 1960
Second Printing, July 1963
Third Printing, August 1967
Fourth Printing, June 1971
Fifth Printing, November 1973
Sixth Printing, April 1976
Seventh Printing, November 1978
Eighth Printing, September 1981
Ninth Printing, August 1984
Plate 1
GEOLOGIC TIME SCALE
William H. Matthews III[1]
Almost everyone has seen the fossilized remains of prehistoric plants or animals. These might have been the skeleton of a gigantic dinosaur, the petrified trunk of an ancient tree, or the shells of snails or oysters that lived in the great seas that covered Texas millions of years ago.
Each year more and more people are learning that these fossils are more than mere curiosities. Instead, they are realizing that a good collection of fossils provides much information about the early history of our earth, and that fossil collecting can be a most enjoyable, fascinating, and rewarding hobby. It is for these people that Texas Fossils was written.
This publication is primarily an amateur collector’s handbook and as such offers many suggestions and aids to those who would pursue the hobby of fossil collecting. It tells, for example, what fossils are, where and how to collect them, and how they are used. Suggestions are made as to how the specimens may be identified and catalogued, and there are discussions and illustrations of the main types of plant and animal fossils. Included also is a simplified geologic map of Texas and a brief review of the geology of the State.
Texas Fossils is not a comprehensive study of the paleontology of Texas. Rather, it deals primarily with the more common species that the average collector is likely to find. These fossils are illustrated in the plates and figures, and these illustrations should be of some help in identifying the specimens in one’s collection. Included for completeness, however, are sketches and descriptions of some of the more rare and unusual fossils, and, for general interest, there are illustrations and descriptions of many of the extinct reptiles and mammals that once inhabited this State.
In addition, a group of selected references has been included for the reader who wishes to know more about earth history and paleontology. Many of these publications provide references of a more technical nature for the more advanced or serious collector, and some of them list excellent collecting localities.
A minimum of technical terminology has been used, but terms not commonly found in dictionaries, or which have not been explained in the text, are defined in the glossary (pp. 111-114).
Many people have helped in the planning, preparation, and completion of Texas Fossils, and their help is gratefully acknowledged: Dr. Keith Young, The University of Texas; Dr. Harold Beaver, Baylor University; and Professor Jack Boon, Arlington State College, offered helpful suggestions and information on Cretaceous fossils; Professors Richmond L. Bronaugh, Baylor University, and Jack T. Hughes, West Texas State College, provided information on vertebrate collecting localities; Professor Fred Smith, Texas A&M College, supplied data on Tertiary collecting localities and fossils which were used in illustrations; Dr. Saul Aronow and Professor Darrell Davis, Lamar State College of Technology; Dr. Jules DuBar, University of Houston; and Dr. Samuel P. Ellison, The University of Texas, made valuable suggestions which have been incorporated into the manuscript.
Special thanks are due Drs. John T. Lonsdale, L. F. Brown, Jr., and Peter U. 2 Rodda, Bureau of Economic Geology, who critically read the manuscript and contributed greatly to the presentation of the material; Dr. John A. Wilson, The University of Texas, who read the section on vertebrate fossils and made invaluable suggestions and criticisms; Miss Josephine Casey, who edited the manuscript; and Mr. J. W. Macon, who prepared the maps and charts.
Thanks are due also to Dr. G. A. Cooper, United States National Museum, who prepared Plate 3 especially for this publication, and to R. T. Bird and the American Museum of Natural History for photographs used in Plates 4 and 43. Plates 38 and 39 were provided through the courtesy of Dr. J. W. Dixon, Jr., and the Geology Department of Baylor University. The other photographs were prepared by the writer. To Sarah Louise Wilson, Lamar State College of Technology, the writer gratefully acknowledges her tireless and painstaking efforts in preparing the many fine drawings which make up the balance of the illustrations.
Fossils are the remains or evidence of ancient plants or animals that have been preserved in the rocks of the earth’s crust. Most fossils represent the preservable hard parts of some prehistoric organism that once lived in the area in which the remains were collected.
The word fossil is derived from the Latin word fossilis, meaning “dug up,” and for many years any unusual object dug out of the ground was considered to be a “fossil.” For this reason some of the earlier books dealing with fossils include discussions of rocks, minerals, and other inorganic objects.
There is much evidence to indicate that man has been interested in fossils since the very earliest times, and fossil shells, bones, and teeth have been found associated with the remains of primitive and prehistoric men. It is quite possible that the owners of these objects believed that they possessed supernatural powers, such as healing properties or the ability to remove curses.
During the earliest periods of recorded history, certain Greek scholars found the remains of fish and sea shells in desert and mountainous regions. These men were greatly puzzled by the occurrence of these objects at such great distances from the sea, and some of them devoted considerable time to an explanation of their presence.
In 450 B.C., Herodotus noticed fossils in the Egyptian desert and correctly concluded that the Mediterranean Sea had once been in that area.
Aristotle in 400 B.C. stated that fossils were organic in origin but that they were embedded in the rocks as a result of mysterious plastic forces at work within the earth. One of his students, Theophrastus (about 350 B.C.), also believed that fossils represented some form of life but thought that they had developed from seeds or eggs that had been planted in the rocks.
Strabo (about 63 B.C. to A.D. 20) was another important Greek scholar who attempted to explain the presence of fossils. He noted the occurrence of marine fossils well above sea level and correctly inferred that the rocks containing them had been subjected to considerable elevation.
During the “Dark Ages” fossils were alternately explained as freaks of nature, the remains of attempts at special creation, and devices of the devil which had been placed in the rocks to lead men astray. These superstitious beliefs and the opposition from religious authorities hindered the study of fossils for hundreds of years.
In approximately the middle of the fifteenth century the true origin of fossils was generally accepted, and they were considered to be the remains of prehistoric organisms which had been preserved in the earth’s crust. With the definite recognition of fossils as organic remains, many of the more primitive theories were discarded for one just as impractical—these remains were considered remnants of the Great Flood as recorded in the Scriptures. The resulting controversy between scientists and theologians lasted for about 300 years.
During the Renaissance several of the early natural scientists concerned themselves with investigations of fossils. Noteworthy among these was Leonardo da Vinci, the famous Italian artist, naturalist, and engineer. Leonardo insisted that the Flood could not be responsible for all fossils nor for their occurrence in the highest mountains. He reaffirmed the belief that fossils were indisputable evidence of ancient life, and that the sea had once covered northern Italy. Leonardo explained that the remains of the animals that had inhabited this ancient body of water were buried in the sediments of the sea floor, and that at some later date in 4 earth history this ocean bottom was elevated well above sea level to form the Italian peninsula.
In the late eighteenth and early nineteenth centuries the study of fossils became firmly established as a science, and since that time fossils have become increasingly important to the geologist.
The study of fossils is called paleontology (Greek palaios, ancient; ontos, a being; logos, word or discourse). Information gathered with the help of paleontology has greatly increased the knowledge of ancient plants and animals and of the world in which they lived.
Fossils represent the remains of such great numbers and various types of organisms that paleontologists have found it helpful to establish four main divisions within their science.
Paleobotany deals with the study of fossil plants and the record of the changes which they have undergone.
This is the study of fossil animals without a backbone or spinal column. These include such forms as fossil protozoans (tiny one-celled animals), snails, clams, starfish, and worms, and usually represent the remains of animals that lived in prehistoric seas.
Because invertebrate remains are the most common fossils in Texas, this book is devoted largely to the discussion of invertebrate fossils and their method of collection.
The vertebrate paleontologist studies the fossils of animals which possessed a backbone or spinal column. The remains of fish, amphibians, reptiles, birds, and mammals are typical vertebrate fossils.
Micropaleontology is the study of fossils that are so small that they are best studied under a microscope. These tiny remains are called microfossils and usually represent the shells or fragments of minute plants or animals. Because of their small size, microfossils can be brought out of wells without being damaged by the mechanics of drilling or coring. For this reason microfossils are particularly valuable to the petroleum geologist who uses them to identify rock formations thousands of feet below the surface.
The majority of fossils are found in marine sedimentary rocks. These are rocks that were formed when salt-water sediments, such as limy muds, sands, or shell beds, were compressed and cemented together to form rocks. Only rarely do fossils occur in igneous and metamorphic rocks. The igneous rocks were once hot and molten and had no life in them, and metamorphic rocks have been so greatly changed or distorted that any fossils that were present in the original rock have usually been destroyed or so altered as to be of little use to the paleontologist.
But even in the sedimentary rocks only a minute fraction of prehistoric plants and animals have left any record of their existence. This is not difficult to understand in view of the rather rigorous requirements of fossilization.
Although a large number of factors ultimately determine whether an organism will be fossilized, the three basic requirements are:
1. The organism should possess hard parts. These might be shell, bone, teeth, or the woody tissue of plants. However, under very favorable conditions of preservation it is possible for even such fragile material as an insect or a jellyfish to become fossilized.
2. The organic remains must escape immediate destruction after death. If the body parts of an organism are crushed, decayed, or badly weathered, this may result in the alteration or complete destruction of the fossil record of that particular organism.
3. Rapid burial in a material capable of retarding decomposition. The type of material burying the remains usually depends upon where the organism lived. The remains of marine animals are common as fossils because they fall to the sea floor after death, and here they are covered by soft muds which will be the shales and limestones of later geologic periods. The finer sediments are less likely to damage the remains, and certain fine-grained Jurassic limestones in Germany have faithfully preserved such delicate specimens as birds, insects, and jellyfishes.
Ash falling from nearby volcanoes has been known to cover entire forests, and some of these fossil forests have been found with the trees still standing and in an excellent state of preservation.
Quicksand and tar are also commonly responsible for the rapid burial of animals. The tar acts as a trap to capture the beasts and as an antiseptic to retard the decomposition of their hard parts. The Rancho La Brea tar pit at Los Angeles, California, is famous for the large number of fossil bones that have been recovered from it. These include such forms as the sabre-tooth cat, giant ground sloths, and other creatures that are now extinct. The remains of certain animals that lived during the Ice Ages have been incorporated into the ice or frozen ground, and some of these frozen remains are famous for their remarkable degree of preservation.
Although untold numbers of organisms have lived on the earth in past ages, only a minute fraction of these have left any record of their existence. Even if the basic requirements of fossilization have been fulfilled, there are still other reasons why some fossils may never be found.
For example, large numbers of fossils have been destroyed by erosion or their hard parts have been dissolved by underground waters. Others were entombed in rocks that were later subjected to great physical change, and fossils enclosed in these rocks are usually so damaged as to be unrecognizable.
Then, too, many fossiliferous rocks cannot be studied because they are covered by water or great thicknesses of sediments, 6 and still others are situated in places that are geographically inaccessible. These and many other problems confront the paleontologist as he attempts to catalog the plants and animals of the past.
The missing pages in the fossil record become more obvious and more numerous in the older rocks of the earth’s crust. This is because the more ancient rocks have had more time to be subjected to physical and chemical change or to be removed by erosion.
There are many different ways in which plants and animals may become fossilized. The method of preservation is usually dependent upon (1) the original composition of the organism, (2) where it lived, and (3) the forces that affected it after death.
Most paleontologists recognize four major types of preservation, each being based upon the composition of the remains or the changes which they have undergone.
This type of fossil is formed only under very special conditions of preservation. To be preserved in this manner, the organism must be buried in a medium capable of retarding decomposition of the soft parts. Materials that have been known to produce this type of fossilization are frozen soil or ice, oil-saturated soils, and amber (fossil resin). It is also possible for organic remains to become so desiccated that a natural mummy is formed. This usually occurs only in arid or desert regions and when the remains have been protected from predators and scavengers.
Probably the best-known examples of preserved soft parts of fossil animals have been discovered in Alaska and Siberia. The frozen tundra of these areas has yielded the remains of large numbers of frozen mammoths—a type of extinct elephant (Pl. 49). Many of these huge beasts have been buried for as long as 25,000 years, and their bodies are exposed as the frozen earth begins to thaw. Some of these giant carcasses have been so well preserved that their flesh has been eaten by dogs and their tusks sold by ivory traders. Many museums display the original hair and skin of these elephants, and some have parts of the flesh and muscle preserved in alcohol.
Original soft parts have also been recovered from oil-saturated soils in eastern Poland. These deposits yielded the well-preserved nose-horn, a foreleg, and part of the skin of an extinct rhinoceros.
The natural mummies of ground sloths have been found in caves and volcanic craters in New Mexico and Arizona. The extremely dry desert atmosphere permitted thorough dehydration of the soft parts before decay set in, and specimens with portions of the original skin, hair, tendons, and claws have been discovered.
One of the more interesting and unusual types of fossilization is preservation in amber. This type of preservation was made possible when ancient insects were trapped in the sticky gum that exuded from certain coniferous trees. With the passing of time this resin hardened, leaving the insect encased in a tomb of amber, and some insects and spiders have been so well preserved that even fine hairs and muscle tissues may be studied under the microscope.
Although the preservation of original soft parts has produced some interesting and spectacular fossils, this type of fossilization is relatively rare, and the paleontologist must usually work with remains that have been preserved in stone.
Almost all plants and animals possess some type of hard parts which are capable of becoming fossilized. Such hard parts may consist of the shell material of clams, oysters, or snails, the teeth or bones of vertebrates, the exoskeletons of crabs, or the woody tissue of plants. These hard parts are composed of various minerals which are capable of resisting weathering and chemical action, and fossils of this sort are relatively common.
Many of the fossil mollusks found in the Tertiary and Cretaceous rocks of Texas have been preserved in this manner. In some of the specimens the original shell material is so well preserved that the iridescent mother-of-pearl layer of the shell is found virtually intact. This type of preservation is less common, however, in the older rocks of the State.
PLATE 2
Types of Fossil Preservation
At certain localities in north and central Texas the Woodbine sands of Upper Cretaceous age (geologic time scale and geologic map, Pls. 1, 10) contain large numbers of shark and fish teeth (Pl. 37), fish scales and vertebrae. The remains of these vertebrates are unusually well preserved and are prized by both amateur and professional collectors.
Hard parts composed of calcite (calcium carbonate) are very common among the invertebrates. This is particularly true of the shells of clams, snails, and corals. Many of these shells have been preserved with little or no evidence of physical change (Pl. 2).
The bones and teeth of vertebrates and the exoskeletons of many invertebrates contain large amounts of calcium phosphate. Because this compound is particularly weather resistant, many phosphatic remains (such as the fish teeth in the Woodbine sands) are found in an excellent state of preservation.
Many organisms having skeletal elements composed of silica (silicon dioxide) have been preserved with little observable change. The siliceous hard parts of many microfossils and certain types of sponges have become fossilized in this manner (Pl. 14).
Some organisms have an exoskeleton (outer body covering) composed of chitin, a material that is similar to finger nails. The fossilized chitinous exoskeletons of arthropods and other organisms are commonly preserved as thin films of carbon because of their chemical composition and method of burial.
The original hard parts of an organism normally undergo great change after burial. These changes take place in many ways, but the type of alteration is usually determined by the composition of the hard parts and where the organism lived. Some of the more common processes of alteration are discussed below.
This process, known also as distillation takes place as organic matter slowly decays after burial. During the process of decomposition, the organic matter gradually loses its gases and liquids leaving only a thin film of carbonaceous material (Pl. 2, fig. 7). This is the same process by which coal is formed, and large numbers of carbonized plant fossils have been found in many coal deposits.
In Texas the carbonized remains of plants, fish, and certain invertebrates have been preserved in this manner, and some of these carbon residues have accurately recorded even the most minute structures of these organisms.
Many fossils have been permineralized or petrified—literally turned to stone. This type of preservation occurs when mineral-bearing ground waters infiltrate porous bone, shell, or plant material. These underground waters deposit their mineral content in the empty spaces of the hard parts making them heavier and more resistant to weathering. Some of the more common minerals deposited in this manner are calcite, silica, and various compounds of iron.
This type of preservation takes place when the original hard parts of organisms are removed after being dissolved by underground water. This is accompanied by almost simultaneous deposition of other substances in the resulting voids. Some replaced fossils will have the original structure destroyed by the replacing minerals. 11 Others, as in the case of certain silicified tree trunks, may be preserved in minute detail.
Although more than 50 minerals have been known to replace original organic structures, the most frequent replacing substances are calcite, dolomite (a calcium magnesium carbonate), silica, and certain iron compounds.
Calcareous replacement occurs when the hard parts of an organism are replaced by calcite, dolomite, or aragonite (a mineral which is composed of calcium carbonate but which is less stable than calcite). The exoskeletons of many corals, echinoderms, brachiopods, and mollusks have been replaced in this manner.
When the original organic hard parts have been replaced by silica the fossil is said to have undergone silicification, and this type of replacement often produces a very high degree of preservation. This is particularly true of the silicified Permian (geologic time scale, Pl. 1) fossils from the Glass Mountains in Brewster County. These fossils are embedded in limestone which must be dissolved in vats of acid, and after the enclosing rock has been dissolved the residue yields an amazing variety of perfectly preserved invertebrate fossils (Pl. 3).
Silicified Cretaceous fossils have been recovered from the Edwards limestone of central Texas. The silicified fauna is restricted to a few scattered localities, each of which may yield many unusually well-preserved fossils.
Several different iron compounds have been known to replace organic matter. Many Texas limestones contain fossil snails and clams which have had their original shell material replaced by iron compounds such as limonite, hematite, marcasite, or pyrite. Certain of the fossiliferous Tertiary sandstones of the Texas Gulf Coast area contain large amounts of glauconite which commonly replaced organic material.
In some areas entire faunas have been replaced by iron compounds. Such is the case in the famous “Pyrite Fossil Zone” of the Pawpaw formation (Lower Cretaceous) in Tarrant County. The fossils in this part of the formation are very small or “dwarfed” and have been replaced by limonite, hematite, or pyrite. Ammonites, clams, snails, and corals are particularly abundant at this locality.
Fossils consist not only of plant and animal remains but of any evidence of their existence. In this type of fossilization there is no direct evidence of the original organism, rather there is some definite indication of the former presence of some ancient plant or animal. Objects of this sort normally furnish considerable information as to the identity or characteristics of the organism responsible for them.
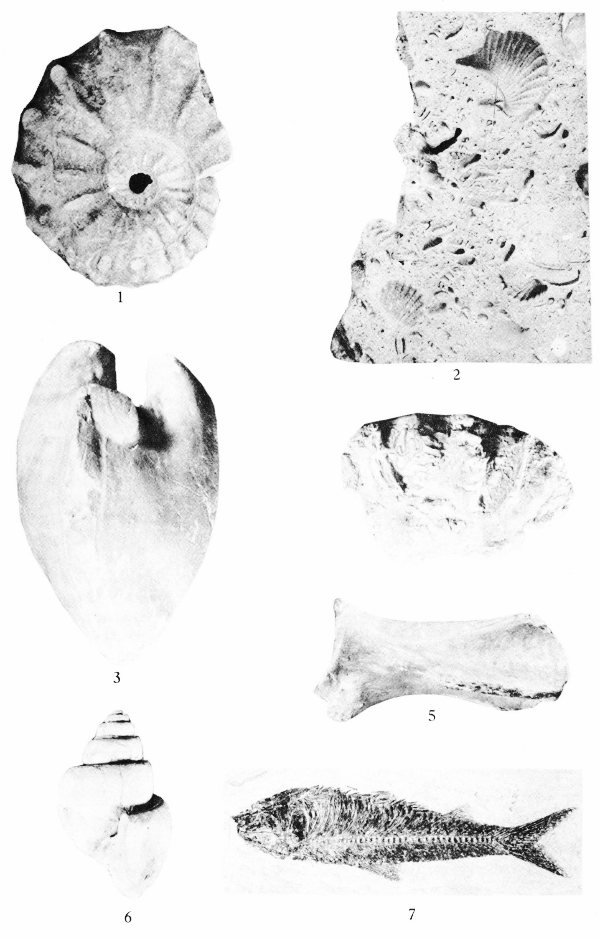
Many shells, bones, leaves, and other forms of organic matter are preserved as molds and casts. If a shell had been pressed down into the ocean bottom before the sediment had hardened into rock, it may have left the impression of the exterior of the shell. This impression is known as a mold (Pl. 2). If at some later time this mold was filled with another material, this produced a cast. This cast will show the original external characteristics of the shell. Such objects are called external molds if they show the external features of the hard parts (Pl. 2, fig. 2) and internal molds (Pl. 2, fig. 3) if the nature of the inner parts is shown.
Molds and casts are to be found in almost all of the fossil-bearing rocks of Texas, and they make up a large part of most fossil collections. It is particularly common to find fossil clams and snails preserved by this method. This is primarily because their shells are composed of minerals that are relatively easy to dissolve, and the original shell material is often destroyed.
PLATE 3
Silicified Brachiopods
All specimens from Permian limestones of the Glass Mountains, Brewster County, Texas

Many animals have left records of their movements over dry land or the sea bottom. Some of these, such as footprints (Pl. 4), indicate not only the type of animal that left them but often provide valuable information about the animal’s environment.
Thus, the study of a series of dinosaur tracks would not only indicate the size and shape of the foot but also provide some information as to the weight and length of the animal. In addition, the type of rock containing the track would help determine the conditions under which the dinosaur lived.
Some of the world’s most famous dinosaur tracks are to be found in the Lower Cretaceous limestones in Somervell County, Texas. These footprints, which are about 110,000,000 years old (Pl. 4), were discovered in the bed of Paluxy Creek near the town of Glen Rose. Large segments of the rock containing these tracks were collected by paleontologists of the American Museum of Natural History in New York City and the Texas Memorial Museum at Austin. Great slabs of limestone were transported to the museums, replaced in their original position, and are now on display as mute evidence of the gigantic size of these tremendous reptiles.
Invertebrates also leave tracks and trails of their activities, and these markings may be seen on the surfaces of many sandstone and limestone deposits. These may be simple tracks, left as the animal moved over the surface, or the burrows of crabs or other burrowing animals. Markings of this sort provide some evidence of the manner of locomotion of these organisms and of the type of environment that they inhabited.
Coprolites are fossil dung or body waste (fig. 1). These objects can provide valuable information as to the food habits or anatomical structure of the animal that made them.

Fig. 1. Sketch of a coprolite—fossilized animal excrement.
These highly polished well-rounded stones (fig. 2) are believed to have been used in the stomachs of reptiles for grinding the food into smaller pieces. Large numbers of these “stomach stones” have been found with the remains of certain types of dinosaurs.
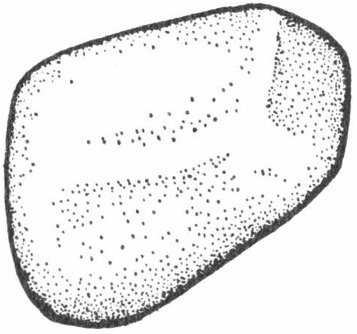
Fig. 2. Sketch of a gastrolith—the gizzard stone of an ancient reptile.
Among the many inorganic objects formed by nature there are some that bear superficial resemblance to plants or animals. Because they are often mistaken for organic remains, these objects have been called pseudofossils, or “false fossils.”
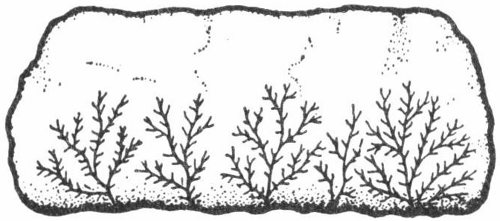
Fig. 3. Dendrites. These thin branching mineral deposits bear a marked resemblance to plants, hence they are called pseudofossils.
Although these closely resemble the remains of ferns or other plant material (fig. 3), dendrites are actually thin incrustations of manganese dioxide. They are often found along the bedding planes of Cretaceous and Paleozoic (geologic time scale, Pl. 1) limestones in many parts of Texas.
Plate 4
Dinosaur tracks in limestone in bed of Paluxy Creek near Glen Rose, Somervell County, Texas.
Photograph courtesy of the American Museum of Natural History.
Permission to reproduce by R. T. Bird.

These are striations that are produced when rock surfaces move past each other while being fractured. Slickensides may superficially resemble certain of the Pennsylvanian coal plants of Texas.
Since slickensides are commonly at an angle to the bedding plane and plant remains lie parallel to the bedding plane, the two are usually easily distinguished.
Many shales and sandstones contain hardened masses of minerals and rock that are often mistaken for fossils. These masses, called concretions, are usually found weathered out of the surrounding rock and may assume the shape of bones, flowers, vegetables, turtles, etc. Although these concretions do not represent organic remains, it is sometimes possible to find true fossils inside them.
In fossil collecting, as in most “collecting” hobbies, the key to success lies in knowing where to look, what equipment to use, and the most effective methods of collecting.
Fossil collecting is a relatively inexpensive hobby because it requires a minimum of supplies and equipment. However, as in almost any hobby, there are certain basic items of equipment that must be acquired.
The hammer is the basic tool in the collector’s kit. Almost any type of hammer is satisfactory, but as collecting experience is gained it may be desirable to get a geologist’s hammer. These hammers, also called mineralogist’s or prospector’s picks, are of two types. One type has a square head on one end and a pick on the other (Pl. 5): the other type is similar to a stonemason’s or bricklayer’s hammer and has a chisel end instead of the pointed pick end. The square head of the hammer is useful in breaking or chipping harder rocks, and the chisel or pick end is good for digging, prying, and splitting soft rocks.
It will be necessary to have some type of bag in which to carry equipment, fossils, and other supplies. A Boy Scout knapsack, musette bag (Pl. 5), hunting bag, or similar canvas or leather bag is suitable.
A pair of chisels is useful when fossils must be chipped out of the surrounding rock. Two sizes, preferably ½ and 1 inch, will usually suffice. A small sharp punch or awl is effective in removing smaller specimens from the softer rocks.
Some specimens are more fragile than others, and these should be handled with special care. Several sheets of newspaper should always be kept in the collecting bag, and each specimen should be wrapped individually as it is collected. Such precautions taken in the field will usually prevent prized specimens from being broken or otherwise damaged. In addition to newspaper, it is wise to carry a supply of tissue paper in which to wrap more fragile specimens.
It is most important to have some method of recording where the fossils were found. It is very easy to forget where the material was collected, and one should never rely on memory. A small pocket-sized notebook is inexpensive and just the right size to carry in the field.
A highway or county map should be used to find the geographic location of each collecting locality. Maps of Texas counties can be obtained from the Texas Highway Department, File D-10, Austin 14, Texas. These maps come in three different sizes, but for most purposes the 18×25-inch sheets, with a scale of ½ inch = 1 mile, will be satisfactory. These are available for all counties and may be purchased at a nominal price.
A magnifying glass or hand lens (Pl. 5) is useful for looking at small specimens and will also prove helpful in examining the finer details of larger fossils. A 10-power magnification is satisfactory for most purposes, and several inexpensive models are available.
Small bags are useful in separating specimens from different localities. Heavy-duty hardware bags for large rough material and medium-weight grocery bags for smaller specimens may be used. Locality data may be written directly on the bag or on a label placed inside with the fossils. As an added precaution some collectors do both. The more serious collector may want to use a cloth geological sample bag (Pl. 5).
Plate 5
FOSSIL COLLECTING EQUIPMENT
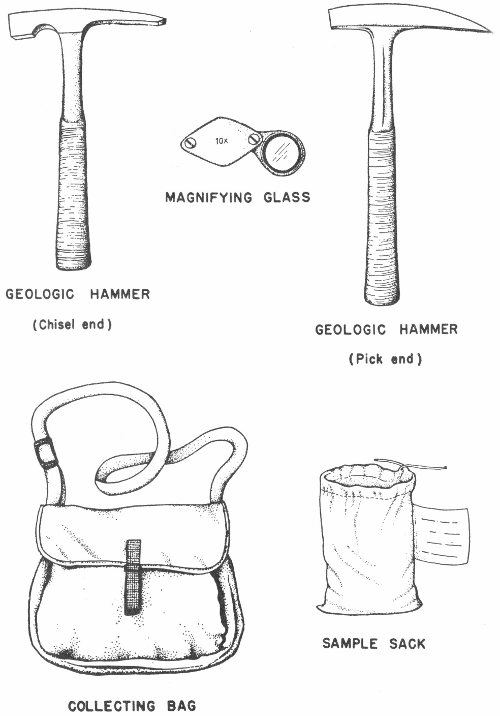
The items described above are those that are most needed and constitute the basic equipment of the fossil hunter. The serious amateur may wish to include certain additional items which will place his collecting on a more professional basis. Some of these accessory items are:
1. A topographic map of the collecting area. These are available for many parts of the State and are published and distributed at nominal cost by the United States Geological Survey, Washington, D. C., and/or Denver, Colorado. The Survey can supply an index sheet showing all such maps available for Texas.
2. A geologic map of the collecting area if one is available. The list of publications of the Bureau of Economic Geology should be consulted to see if a geologic report or map of the area has been published. This list may be obtained without charge from the Bureau of Economic Geology, The University of Texas, Austin 12, Texas.
3. The geologic map of Texas. Although a geologic map of Texas is included in this publication (Pl. 10), the scale is so small that its use is somewhat limited. For more detailed work a larger geologic map in color (scale: 1 inch = 31.56 miles) may be ordered from the Bureau. The sale price is 25 cents.
4. A compass for more accurate location of collecting localities.
5. Adhesive or masking tape. The locality information can be written on the tape and applied directly to the specimen.
6. Paper labels (about 3×5 inches). A properly completed label should be placed inside each bag of material.
Knowing where to look for fossils is a very important part of fossil collecting. It has already been pointed out that igneous and metamorphic rocks are not likely to be fossiliferous, but that most fossils are found in marine sedimentary rocks. These sediments were deposited under conditions that were favorable for organisms during life and which facilitated preservation after death. Limestones, limy shales, and certain types of sandstones are typically deposited under such conditions.
One should look particularly for areas where rocks formed from marine sediments lie relatively flat and have not been greatly disturbed by heat, pressure, and other physical or chemical changes. If the rocks appear to have undergone considerable folding and fracturing, there is great likelihood that any fossils that were present have been destroyed or damaged by this action.
Quarries are good places to look but one should be sure to obtain permission before entering. Rock exposures in quarries are rather fresh but have undergone some weathering. Quarries have been opened in many of the limestone formations of Texas, and large numbers of fine specimens have been collected in some of these excavations. Certain Lower Cretaceous limestones are useful for road metal, building stone, or in the manufacture of portland cement, and extensive quarrying has been undertaken in the Edwards Plateau region of Texas (Pl. 9). Bones and petrified wood are frequently found in sand and gravel quarries in many parts of the State.
Particular attention should be given to all railroad and highway cuts as rocks exposed in this way are usually still in their original position and are fairly well weathered. Cuts made by recent construction are usually more productive after they have undergone a period of weathering as this helps to separate the fossils from their enclosing rocks.
Gullies, canyons, and stream beds are also good places to examine. These areas are continually subjected to the processes 20 of erosion or stream action, and new material is uncovered year after year.
If there are abandoned coal mines nearby, the dumps of waste rock around the mine shafts could be checked. A careful examination of such waste may reveal fine specimens of well-preserved plant fossils.
Coal has been mined in several parts of Texas, and abandoned shafts or dumps are still present in some counties. The bituminous coals of Texas are predominantly Pennsylvanian in age, and mining has been carried on in the following counties: Eastland, Erath, Jack, Palo Pinto, Parker, Wise, Young.
When a likely collecting spot has been located, the ground should be examined very carefully to see if there are any rock fragments which contain pieces of shell or the imprints of leaves or other organisms.
If the fossils have been freed by weathering, they can be easily picked up and placed in the bag. Many times, however, it will be necessary to take the hammer and very carefully remove the surrounding rock. Smaller specimens may be more safely freed with the careful use of the proper size chisel by gently tapping the chisel and gradually chipping away the matrix—the rock that is holding the specimen. After most of the matrix has been removed, the fossil should be carefully wrapped and placed in the collecting bag.
Before leaving a collecting locality, one should be sure to record its geographic location and the geologic age of the rock in which the fossils were found. The place should be located on the map and the locality entered in the notebook in such a manner that it could easily be located again for additional collecting. If a county or topographic map is available, it is wise to mark the locality on the map. The geographic and geologic data should be written on a label placed in the bag of fossils collected at that particular locality. In addition, many collectors find it helpful to write the locality on the outside of each bag of fossils.
Material from separate localities should be kept in individual cloth or paper bags, and the collector should take every precaution to keep the labels with their respective fossils. Remember that a fossil without a locality is hardly worth the paper it is wrapped in.
The collector should always ask the land owner’s permission before entering or collecting on private property. One should respect all property, especially livestock and fences, and leave the area cleaner than when entered. If these precautions are observed, future collectors will probably be welcome to return for additional collecting.
It is usually necessary to do the final cleaning and preparation of fossils at home or in the laboratory, for most fossils brought in from the field require considerable preparation before they are ready for display.
Excess matrix should be carefully removed with hammer and chisel; blows should always be directed away from the fossil. Smaller tools (needles, tweezers, and awls) should be used in the final preparation stage, and one should work carefully to avoid damaging the specimen. Before starting the final cleaning, it will be helpful to place the fossils in water and let them soak overnight. This will loosen much of the excess rock, and most of the softer material can then be removed with a small scrub brush or tooth brush. Mounted needles can be used to clean more delicate specimens or around the smaller structures of larger fossils. It may be advisable to use the magnifying glass when working with small fossils or with delicate surface structures of larger specimens.
Broken fossils can be repaired with clear plastic household cement, and specimens that are crumbling may be coated with pure white shellac, thinned collodion, or clear nail polish. The latter is preferred as it is not as likely to crack. Fragments of bone are particularly apt to crumble upon exposure to the air. This type of fossil is normally quite fragile and should be excavated with great care and shellaced as soon as dry.
Dilute hydrochloric acid may be used in removing silicified fossils from a calcareous matrix. The material to be etched should be placed in a pottery or glass container and covered with water. Acid should then be added to the water very slowly and until large numbers of bubbles are given off. Each time the bubbling ceases, more acid should be added and this process should be repeated until the fossil is free of matrix. This procedure should be carried on in a well-ventilated place, and the acid should be handled with extreme caution. Hydrochloric acid can cause damage or serious injury and the fumes are extremely corrosive.
In order to get the maximum pleasure out of fossil collecting, most amateur paleontologists want to identify and classify the fossils that they have collected. This requires some knowledge of how fossils are classified and how they receive their scientific names.
The number of organisms, both living and extinct, is so great that some system of classification is needed to link them all together. Many fossils bear distinct similarities to plants and animals that are living today, and for this reason paleontological classification is similar to that used to classify modern organisms. This system, known as the system of binomial nomenclature, was first used consistently in 1758 by Linné (or Linnaeus), an early Swedish naturalist.
Scientific names established in accordance with the principles of binomial nomenclature consist of two parts: the generic (or genus) name and the trivial name. These names are commonly derived from Greek or Latin words which are usually descriptive of the organism or fossil being named. They may, however, be derived from the names of people or places, and in such instances the names are always Latinized. Greek or Latin is used because they are “dead” languages and not subject to change. They are also “international” languages in that scientists all over the world can use the same names regardless 22 of what language they write in. The system of binomial nomenclature has led to the development of the science of taxonomy, the systematic classification and naming of plants and animals according to their relationships.
The world of organic life has been divided into the plant and animal kingdoms. These kingdoms have been further divided into larger divisions called phyla (from the Greek word phylon, a race). Each phylum is composed of organisms with certain characteristics in common. For example, all animals with a spinal cord (or notochord) are assigned to the phylum Chordata.
The phylum is reduced to smaller divisions called classes, classes are divided into orders, orders into families, families into genera, and each genus is divided into still smaller units called species. A species may be further reduced to subspecies, varieties, or other subspecific categories, but these need not concern us in a publication of this nature.
The following table illustrates the use of binomial nomenclature in the classification of man, a clam, and a dog.
| Unit | Man | Dog | Clam |
| Kingdom | Animalia | Animalia | Animalia |
| Phylum | Chordata | Chordata | Mollusca |
| Class | Mammalia | Mammalia | Pelecypoda |
| Order | Primates | Carnivora | Eulamellibranchia |
| Family | Hominidae | Canidae | Veneridae |
| Genus | Homo | Canis | Venus |
| Species | sapiens | familiaris | mercenaria |
The generic name and the trivial name constitute the scientific name of a species and according to this system of classification the scientific name of all living men is Homo sapiens. It is obvious that there are many variations among individual men, but all men have certain general characteristics in common and are therefore placed in the same species.
In a scientific name, the generic name is always started with a capital letter and the trivial name with a small letter. Both names must be italicized or underlined.
The name of the author (the person who first described the fossil) usually appears following the scientific name. The date of the scientific publication containing the original description of the fossil is often placed after the author. For example:
Turrilites worthensis Adkins and Winton 1920
With the large numbers of plants and animals that are living today, plus those of the past, random naming would result in much confusion. For this reason scientists have established strict rules that must be followed when a specimen is named. The strict application of these rules enables scientists in all parts of the world to assign scientific names without fear of duplication.
The beginning collector is usually content to know if his specimen is a clam or a snail or a fern or a palm leaf. But as the collection grows, it becomes increasingly desirable to know the scientific name of each fossil.
When he starts to identify fossils it may be helpful to show them to a geology teacher if a college or university is nearby. Most teachers are glad to be of help and will probably have similar specimens in their own collections. As all colleges do not have geology departments, a list of institutions with geologists on their faculties is included at the end of this section of the handbook (p. 27). In addition, many of the science teachers in the public schools are familiar with fossils and can give helpful suggestions as to how to classify material.
Museums are also good places from which to get help. If the museum has a geological collection, it will be most helpful to compare specimens with the fossils in their collections and to ask the museum personnel for advice. In addition to the above sources of information, local professional geologists are usually familiar with the geology of the local area and the paleontological literature of the region.
Possibly local librarians can recommend books, encyclopedias, or other publications that will be of help. Members of a local rock and mineral club, if one is available, are another source of information. Many times these collectors can pass along good ideas and tell exactly which books to consult.
After books or journals describing the fossils of the area have been located, the collected specimens should be closely compared with any illustrations that are shown. Each fossil should be examined carefully, its more characteristic features noted, and it should again be compared with the illustrations and descriptions in the book. The phylum or class to which the specimen belongs should be determined first. For example, the genus and species of a certain fossil may not be known, but it looks like a snail and accordingly it is named a gastropod (for class Gastropoda, the snail class), and this is, at least, a start in determining the scientific name of that particular fossil. The descriptive material in the text of each reference will usually point out the more detailed features which will be diagnostic of the genus or species.
The illustrations and descriptive material in this publication will also be of considerable help in identification. Many illustrations of the more common invertebrate fossils have been included, but the publication was not designed primarily for use in fossil identification. Rather, it is intended to guide the amateur or student who is interested in fossil collecting, and to furnish suggestions as to how collecting may be more effectively pursued.
Fossil identification keys may be useful in helping the beginning collector identify specimens. The collector compares a fossil with the key description and eliminates those characters that do not fit the specimen.
The key used in this handbook is based primarily on symmetry—the orderly arrangement of the parts of an object with reference to lines, planes, or points. The shape of the shell or body, presence or absence of coiling, and presence or absence of body partitions are also useful criteria in identifying fossils. To use the 24 key the beginner should know something about symmetry. Two major types of symmetry are used in this key.
1. Radial symmetry—the symmetrical repetition of parts around an axis. This is the symmetry of a wheel, and any vertical section through the center of the object divides it into symmetrical halves (fig. 4a).
2. Bilateral symmetry—the symmetrical duplication of parts on each side of a plane (fig. 5). The plane divides the object into two halves that are mirror images of each other. This is the symmetry of a plank.
It should be noted that many objects may have both kinds of symmetry. For example: A cone when viewed from the top has radial symmetry and when viewed from the side shows bilateral symmetry (fig. 4a, b).

Fig. 5. Bilateral symmetry as displayed by a typical fossil brachiopod.
An illustration of the use of the key on pages 26-27 follows. Assuming that a specimen displays radial symmetry, this means that it belongs under Part I on the key. If the fossil has a tapering, cylindrical, cone-shaped shell (“A” on the key), the subheadings under the “A” part of the key are examined. Should the specimen have a shell which is round, tapering at one end, with transverse septa or sutures (number 2 under “A”), it is probably a cephalopod. This is indicated on the right hand side of the page. Number 1 under “A” is eliminated because the fossil did not have longitudinal radial partitions within the shell.
Some fossils display no apparent symmetry and such a fossil would be referred to Part III of the key. If this fossil had internal transverse partitions “A” would be eliminated. If the fossil was not a coiled fossil “B” would also be eliminated and we would proceed directly to “C”—uncoiled fossils. If the specimen is a branching twig-like fossil, numbers 1, 2, and 3 would be eliminated and the specimen referred to number 4 (Branching twig-like fossils). Should the specimen have evenly distributed relatively large openings with radial longitudinal partitions or septa, the specimen is probably a colonial coral (“b” under number 4 on the key). The “a” part of number 4 would be eliminated because the coral had large openings and radial longitudinal septa.
Once a tentative identification has been made from the key, pictures and descriptions of this fossil group are examined to establish a more precise identification. It should be remembered that keys are not perfect, and the collector should not expect to be able to identify every specimen with this key.
(Instructions on pages 23-25 for use of key)
Plate 6
Fossil Identification Chart
I RADIAL SYMMETRY

After the fossils have been cleaned and tentatively identified, they should be cataloged. This is necessary to enable the collector to have a record of his collection and to furnish as much information as possible about each individual fossil.
The collecting data can be taken from the labels that were placed in each bag of fossils as they were collected, or from the field notebook. Actually, it is wise to check one against the other. This information should then be entered in some type of record book and also placed on a more permanent label which is put in the tray or box with the fossil. The catalog and label should contain such pertinent data as (1) the scientific name of the fossil, (2) the geologic formation from which the specimen was collected, (3) the exact geographic location of the collecting locality, (4) the name of the collector, (5) the date the fossil was collected, and (6) the catalog number of the specimen. The latter is usually placed in the upper right hand corner of the label (fig. 6) and corresponds with a like number in the record book.

Fig. 6. A brachiopod showing the catalog number on it, and the accompanying label that pertains to the specimen.
The entries in the catalog should be numbered consecutively, and all specimens from the same locality should bear the same number. This number should be written on the fossil with India ink, preferably on any remaining matrix or on some inconspicuous part of the specimen (fig. 6). If the surface of the fossil is too coarse or porous for ink, the catalog number can be written on a small patch of white enamel or clear nail polish painted on the specimen. After the ink has dried it should be coated with a dab of clear shellac or clear nail polish to help preserve the number. If each specimen is numbered, it can easily be identified even if it should become separated from its label.
Fossils are useful in a number of different ways, for each specimen provides some information about when it lived, where it lived, and how it lived.
Fossils are very important, for example, in tracing the development of the plants and animals of our earth. This is possible because the fossils in the older rocks are usually primitive and relatively simple; but a study of similar specimens that lived in later geologic time shows that the fossils become progressively more complex and more advanced in the younger rocks.
Some fossils, for example, the reef-building corals, appear to have always lived under much the same conditions as they live today. Hence, it is reasonably certain that the rocks containing fossil reef corals 32 found in place (that is, where they were originally buried), were deposited in warm, fairly shallow, salt water. By studying the occurrence and distribution of such marine fossils, it is possible to outline the location and extent of prehistoric seas. Moreover, the type of fossils present will frequently give some indication as to the bottom conditions, depth, temperature, and salinity of these ancient bodies of water.
Probably the most important use of fossils is for purposes of correlation—the process of demonstrating that certain rock layers are closely related to each other. By correlating or “matching” the beds containing specific fossils, it is possible to determine the distribution of geologic units of similar age. Some fossils have a very limited vertical or geologic range and a wide horizontal or geographic range. In other words, they lived but a relatively short period in geologic time but were rather widely distributed during their relatively short life. Such fossils are known as index fossils or guide fossils and are especially useful in correlation because they are normally only associated with rocks of one certain age.

Fig. 7. Sketches of two types of micropaleontological slides. (a) Multiple space faunal slide. (b) Single-hole slide.
Microfossils are often very valuable as guide fossils for the petroleum geologist. The micropaleontologist washes the well cuttings from the drill hole and separates the tiny fossils from the surrounding rocks. The specimens are then mounted on special slides (fig. 7) and studied under the microscope. Information derived from these fossils often provides valuable data on the age of the subsurface formation and the possibilities of oil production. Microfossils are particularly valuable in the oil fields of the Gulf Coast region of Texas. In fact, some of the oil-producing zones in this area have even been named for certain key genera of microfossils. For example, the “het” zone of Oligocene age (geologic time scale, Pl. 1) is named for the genus Heterostegina, which is a tiny one-celled animal. Other microfossils, such as fusulinids, ostracodes, spores, and pollens, are also used to identify subsurface formations in many other parts of the State.
Plant fossils are very useful as climatic indicators but are not too reliable for purposes of age determination. They do, however, provide much information about the development of plants throughout geologic time.
The geologic history of our earth has been recorded primarily in marine sedimentary rocks, and this record indicates that our earth is very old and that life has been present for many millions of years. The earth is not only extremely old (more than 3½ billion years of age), but it has also undergone many changes which have taken place slowly but steadily and have greatly affected both the earth and its inhabitants. The earth’s physical features have not always been as they are seen today. Geologic research has shown that mountains now occupy the sites of ancient seas, and that coal is being mined where swamps existed millions of years ago. Furthermore, there is much evidence to indicate that plants and animals have also undergone great change. The trend of this organic change is, in general, toward more complex and advanced forms of life, but some forms have remained virtually unchanged and others have become extinct.
In order to interpret geologic history, the earth scientist must attempt to gather evidence of the great changes in climate, geography, and life that took place in the geologic past. The record of these changes can be found in the rocks, and here is found the story of the various events in earth history.
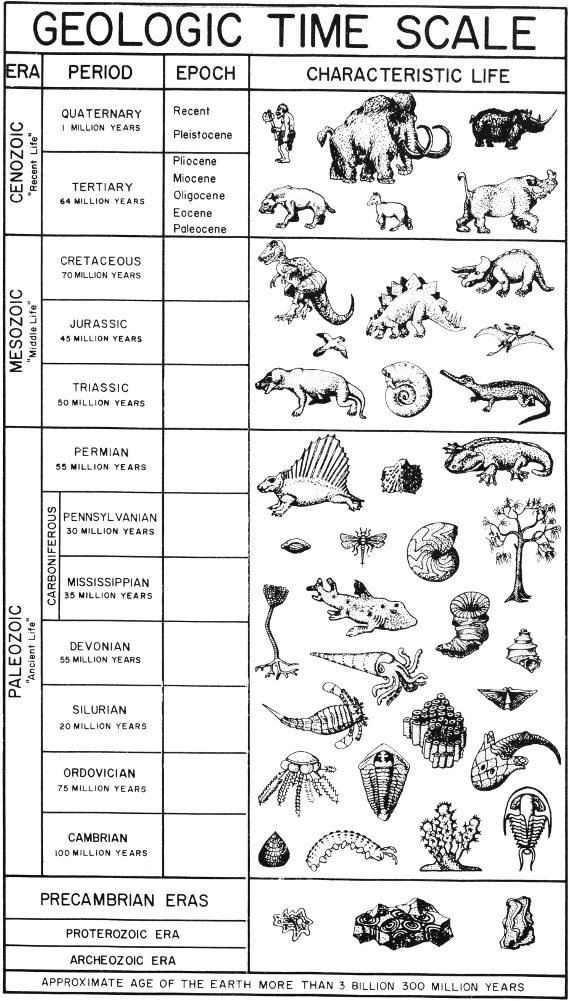
In order to discuss fossils and the age of the rocks containing them, it is necessary to become familiar with the geologic column and the geologic time scale (Pl. 1).
The geologic column refers to the total succession of rocks, from the oldest to most recent, that are found either locally or in the entire earth. Thus, the geologic column of Texas includes all rock divisions known to be present in this State. By referring to the geologic column previously worked out for any given area, the geologist can determine what type of rocks he might expect to find in that particular region.
The geologic time scale is composed of units which represent intervals of geologic time, during which were deposited the rocks represented in the geologic column. These time units are used by the geologist to date the events that have taken place in the geologic past.
The largest unit of geologic time is an era, and each era is divided into smaller time units called periods. A period of geologic time is divided into epochs, which, in turn, may be subdivided into still smaller units. The geologic time scale might be roughly compared to the calendar in which the year is divided into months, months into weeks, and weeks into days. Unlike years, however, geologic time units are arbitrary and of unequal duration, and the geologist cannot be positive about the exact length of time involved in each unit. The time scale does, however, provide a standard by which he can discuss the age of fossils and their surrounding rocks. By referring to the time scale it may be possible, for instance, to state that a certain event occurred during the Paleozoic era in the same sense that one might say that something happened during the American Revolution.
There are five eras of geologic time, and each has been given a name that is descriptive of the degree of life development that characterizes that era. Hence, Paleozoic means “ancient-life,” and the era was so named because of the relatively simple and ancient stage of life development.
The eras, a guide to their pronunciation, and the literal translation of each name is shown below.
Archeozoic and Proterozoic rocks are commonly grouped together and referred 34 to as Precambrian in age. The Precambrian rocks have been greatly contorted and metamorphosed, and the record of this portion of earth history is most difficult to interpret. Precambrian time represents that portion of geologic time from the beginning of earth history until the deposition of the earliest fossiliferous Cambrian strata. If the earth is as old as is believed, Precambrian time may represent as much as 85 percent of all geologic time.
The oldest era is at the bottom of the list because this part of geologic time transpired first and was then followed by the successively younger eras which are placed above it. Therefore, the geologic time scale is always read from the bottom of the chart upward. This is, of course, the order in which the various portions of geologic time occurred and during which the corresponding rocks were formed.
As mentioned above, each of the eras has been divided into periods, and most of these periods derive their names from the regions in which the rocks of each were first studied. For example, the Pennsylvanian rocks of North America were first studied in the State of Pennsylvania.
The Paleozoic era has been divided into seven periods of geologic time. With the oldest at the bottom of the list, these periods and the source of their names are:
The Carboniferous period in Europe includes the Mississippian and Pennsylvanian periods of North America. Although this classification is no longer used in the United States, the term Carboniferous will be found in many of the earlier geological publications and on many of the earlier geologic maps.
The periods of the Mesozoic era and the source of their names are:
In Texas, the Cretaceous has two divisions, known as either Lower Cretaceous and Upper Cretaceous or as Comanche series and Gulf series, respectively. These designations are for rocks of nearly equivalent age, and both sets of terms have been used by geologists and in publications. In this handbook, both sets of terms are used interchangeably, that is, Lower Cretaceous and/or Comanche series and Upper Cretaceous and/or Gulf series.
The Cenozoic periods derived their names from an old outdated system of classification which divided all of the earth’s rocks into four groups. The two divisions listed below are the only names of this system which are still in use:
While the units discussed above are the major divisions of geologic time, the geologist usually works with smaller units of rocks called formations. A geologic formation is identified and established on the basis of definite physical and chemical characteristics of the rocks. Formations are usually given geographic names which are combined with the type of rock that makes up the bulk of the formation. For example, the Beaumont clay was named from clay deposits that are found in and around Beaumont, Texas.
The geologic history of Texas, like the geologic history of the rest of the earth, is recorded primarily in marine sedimentary rocks. These rocks provide some knowledge of the early geography and the first inhabitants of what is now the State of Texas. Most of these rocks were formed 35 from sediments deposited in shallow seas which covered parts of the State at various times in earth history.
By studying these rocks and their relations to each other, geologists have established a geologic column for Texas.
In order to discuss the distribution and exposures of the rocks of Texas, it is helpful to be familiar with the physiography of the State. Physiography deals with the study of the origin and description of land forms, such as mountains, valleys, and plains. Plate 9 is a map of Texas which shows the major physiographic provinces within the State.
The majority of the land forms in Texas have been produced by the processes of erosion attacking the structural features of an area. Certain other land forms may be related to the effects of igneous activity which resulted in the accumulation of large masses of igneous rocks. The Davis Mountains are an example of surface features produced in this manner.
In discussing the physiography of Texas, three major physiographic provinces will be recognized. These are (1) the Trans-Pecos region, (2) the Texas Plains, and (3) the Gulf Coastal Plain (Pl. 9).
The Trans-Pecos region, located in the westernmost part of the State, is an area of mountains and plateaus with broad basins between the major mountain ranges. Many different types of rocks are exposed in Trans-Pecos Texas and these include marine, fresh-water, and terrestrial deposits. In many areas igneous rocks flowed out on the surface and now overlie sedimentary rocks. There are also many places where igneous rocks have been injected into the surrounding rocks, and these igneous rocks have been exposed by later erosion.
Included within this area is the Van Horn uplift of southern Hudspeth and Culberson counties, the Solitario uplift of southern Presidio and Brewster counties, and the Marathon uplift of northeast Brewster County. This region also includes the Big Bend area of Texas, a part of which has been set aside as a National Park where many interesting and important geological features may be seen.
The Trans-Pecos region is one of rugged topography with elevations as high as 8,700 feet, at Guadalupe Peak in the Guadalupe Mountains of northern Culberson County, and as low as 1,500 feet, in the Rio Grande valley.
Numerous invertebrate fossils occur in the Cretaceous limestones and shales of the Trans-Pecos region and in the Paleozoic rocks of the Marathon uplift. The Gaptank formation of Pennsylvanian age and the Permian reef limestones of the Glass Mountains are especially fossiliferous. In addition, many vertebrate fossils have been collected in Trans-Pecos Texas, particularly in and around Big Bend National Park.
The plains of Texas are broad expanses of country with very little surface relief. Most of the plains support grasses and some have wooded areas, particularly along stream valleys.
The plains of the northwestern part of the State have been subdivided as follows.
This area (Pl. 9), often called “the caprock,” is an elevated plateau which rises above the rolling plains which surround it. The High Plains are bounded by the Pecos River valley on the south, southeast, and west and by the North-Central Plains on the east.
The surface of the High Plains is very flat and characterized by a sparse cover of grasses and few trees. The surface strata consist largely of unconsolidated deposits of sands and gravels of Quaternary and Tertiary age, with remnants of Lower Cretaceous limestones along the southern margin. The rocks of the High Plains are mostly unfossiliferous, but mammalian remains have been found at several localities.
Plate 9
Physiographic map of Texas.
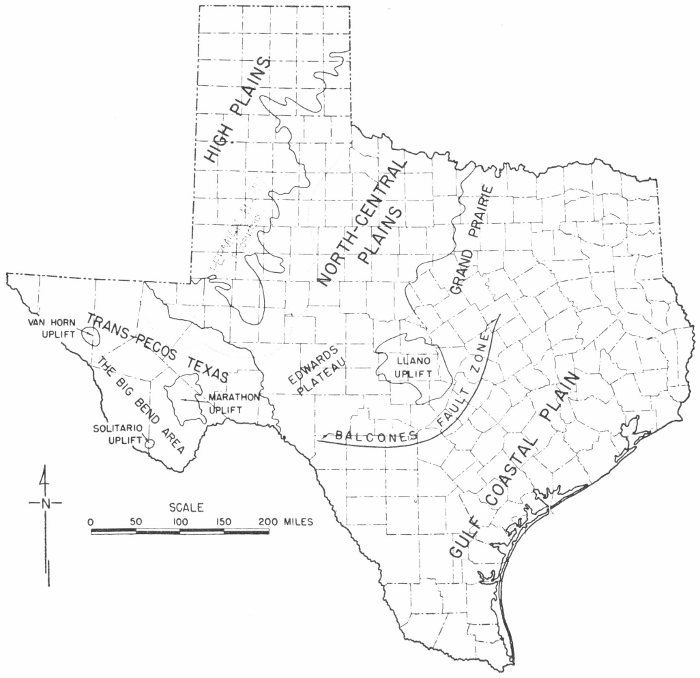
Surface strata of the North-Central Plains (Pl. 9) are westward-dipping Pennsylvanian, Permian, and Triassic rocks. Present also are extensive exposures of Quaternary sands and gravels which trend north-south across the central portion of the region. The area is bounded on the west by the High Plains, on the east by the Grand Prairie, and on the south by the Edwards Plateau and Llano uplift. Many vertebrate fossils have been collected from the Permian and Triassic rocks of this area. There are also many excellent outcrops of fossiliferous Pennsylvanian formations in the North-Central Plains region.
The Edwards Plateau (Pl. 9) is located in south-central Texas and is bounded on the south by the Balcones fault zone and on the north by the North-Central Plains. The surface of the area is typically flat with a gentle slope to the south. The rocks of the Edwards Plateau consist primarily of Lower Cretaceous limestones and shales, many of which are very fossiliferous.
This area (Pl. 9) has a relatively flat surface but there are areas of gently rolling hills. The eastern boundary of the Grand Prairie is marked partly by the Balcones fault zone. North of McLennan County, however, the Balcones fault zone is not expressed at the surface and in this area the eastern boundary is defined by the western edge of the Woodbine exposures. Upper and Lower Cretaceous rocks occur at the surface and dip to the southeast; many of these rocks contain a large number of invertebrate fossils.
The Llano uplift (Pl. 9) is located in the central part of the State where Precambrian igneous and metamorphic rocks and sedimentary rocks of early Paleozoic age occur on the surface. The area, which now appears as a basin-shaped depression, was at one time covered by Lower Cretaceous rocks and perhaps also by Devonian, Mississippian, and Pennsylvanian strata. These have since been removed by erosion. The east, south, and west sides of the uplift are surrounded by Lower Cretaceous rocks, and the northern margin is marked by the Mississippian and Pennsylvanian formations of the North-Central Plains. The area is, in general, composed of unfossiliferous rocks, but some invertebrate fossils (primarily trilobites and brachiopods) have been collected.
The Gulf Coastal Plain (Pl. 9) is composed of Cretaceous, Tertiary, and Quaternary rocks and includes the eastern, southeastern, and southern portions of the State. The rocks of the area consist of sands, clays, shales, and limestones. The Texas Gulf Coastal Plain is bounded on the north and west by the Balcones fault zone, on the south and southwest by the Gulf of Mexico, and extends eastward into Arkansas and Louisiana.
The region has broad river valleys and uplands of low relief, but there is an increase in relief toward the interior of the State. The surface of the area slopes gradually toward the Gulf and successively younger formations are encountered gulfward.
The rocks of the Texas Gulf Coastal Plain are relatively unfossiliferous, but many of the Upper Cretaceous rocks contain fossils. In the central portion of the region some marine formations of Tertiary age locally contain well-preserved invertebrate fossils.
Geologic studies of the State of Texas have indicated the presence of rocks formed during every era and period of geologic time. These range from the Precambrian granites of the Llano uplift to the Quaternary gravels of the High Plains.
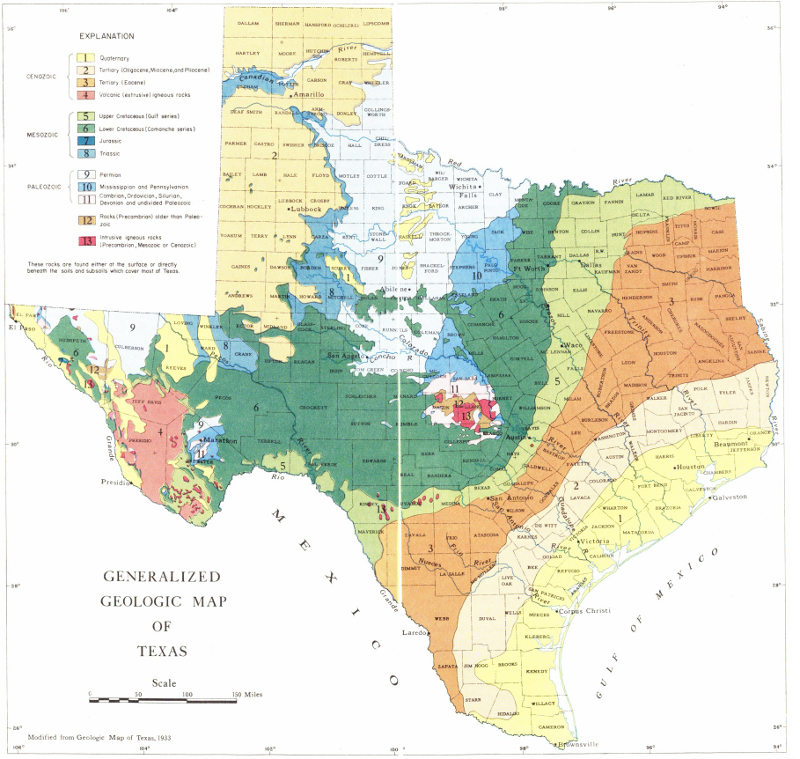
Plate 10
GENERALIZED GEOLOGIC MAP OF TEXAS
Modified from Geologic Map of Texas, 1933
One of the best ways to become acquainted with the geology of Texas is to study the geologic map of the State (Pl. 10). A geologic map shows the distribution and age of surface rocks and may also indicate what kind of geologic structures are present. The types of rocks that crop out at the surface may be shown by means of symbols, colors, or patterns, and these are explained by a legend which accompanies the map. On Plate 10, colors are used to show the distribution and geologic age of the surface rocks of Texas. Reference to this map will give the collector some idea of the age of the fossils that might be found in a given area. Some special geologic maps may have the location of geologic structures and formation contacts indicated by means of symbols, such as dashed lines, arrows, and similar special markings. However, the map included in this publication does not show any of these special markings.
The Precambrian rocks of Texas are composed of igneous and metamorphic rocks and some sedimentary rocks. Most of the Precambrian outcrops are in the Llano uplift and El Paso and Van Horn regions.
Alterations produced by vast amounts of time, heat, and pressure have obliterated any trace of fossils that may have been present in these rocks. With the exception of some questionable primitive plants collected in the Van Horn region, no Precambrian fossils have been reported from Texas.
Rocks of Paleozoic age are widespread in Texas, and rocks of each period are well exposed. Outcrops are found in the Llano uplift, North-Central Plains, and Trans-Pecos region. The most extensive exposures are of Pennsylvanian and Permian age, and the former are highly fossiliferous in parts of the North-Central Plains.
Rocks of late Cambrian age are exposed in the Llano, Marathon, and Solitario uplifts, and the Franklin Mountains near El Paso. These are sedimentary rocks consisting of conglomerates, sandstones, shales, limestones, and some dolomites.
Some of these formations are relatively fossiliferous, but the specimens are commonly fragmental and very poorly preserved. Fossils that are apt to be found in the Cambrian rocks of the Llano uplift include brachiopods, gastropods, trilobites, and small rounded objects believed to have been formed by algae (primitive one-celled plants). In other parts of the State, Cambrian rocks are sparsely fossiliferous and the fossils consist primarily of fragmental brachiopods, trilobites, and algae.
Ordovician outcrops are present in the Llano uplift of central Texas and in the Marathon, Solitario, El Paso, and Van Horn regions of Trans-Pecos Texas. These are sedimentary rocks and consist largely of sandstones, cherts, limestones, and dolomites.
Although some of the Ordovician formations are fossiliferous, they are seldom collected by amateur paleontologists because they are exposed in relatively inaccessible places and the fossils are usually poorly preserved. Ordovician fossils reported from Texas include sponges, corals, brachiopods, gastropods, cephalopods, and trilobites. In addition, the Marathon formation of the Marathon uplift contains large numbers of well-preserved graptolites (fig. 24, p. 86).
The Silurian of Texas is poorly represented in surface exposures, and only one formation, the Fusselman, has been described. The Fusselman crops out in the El Paso and Van Horn regions where it is a white dolomitic limestone. Fossils are not abundant in this formation, but brachiopods and corals have been collected at a few localities.
Devonian rocks are best developed in Trans-Pecos Texas, especially in the Marathon, 41 El Paso, and Van Horn regions. In addition to the Trans-Pecos exposures, there are minor outcrops of Devonian rocks in the Llano uplift of central Texas.
Fossils are rare and fragmental in the Trans-Pecos exposures and consist primarily of radiolarians and brachiopods. The Devonian rocks of central Texas are predominantly calcareous and, although the material is usually poorly preserved, many fossils have been collected from them. These include bryozoans, corals, brachiopods, gastropods, and trilobites. Conodonts and fragments of primitive armored fishes (Pl. 37) have also been reported.
Mississippian rocks are exposed in the Llano region and in the Hueco Mountains of the Trans-Pecos area. The Trans-Pecos rocks primarily contain brachiopods with some bryozoans and gastropods.
The central Texas Mississippian rocks are much more fossiliferous and some of the material is well preserved. Fossils reported from this area include brachiopods (Pl. 17), crinoids, gastropods, cephalopods, trilobites, and ostracodes.
Pennsylvanian rocks are well represented in Texas and are exposed in the Llano uplift, north-central Texas, and Trans-Pecos Texas.
In Trans-Pecos Texas fossiliferous rocks crop out in the Hueco and Diablo Mountains. Fossils found in this area are algae, fusulinids, corals, brachiopods, pelecypods, gastropods, cephalopods, and crinoids. There is also a thick section of Pennsylvanian rocks in the Marathon uplift, but only one formation, the Gaptank, is very fossiliferous. It contains many fossils including fusulinids, sponges, corals, bryozoans, brachiopods, gastropods, pelecypods, cephalopods, and crinoids.
Certain Pennsylvanian strata in the Llano region are very fossiliferous, and the material is well preserved. The more abundant forms are fusulinids, corals, brachiopods, gastropods, pelecypods, cephalopods, and crinoids.
Probably the best Pennsylvanian collecting areas are to be found in north-central Texas. Here the thick marine limestones and shales contain large numbers of well-preserved invertebrate fossils, and the terrestrial or shallow marine strata have yielded an abundance of plant fossils. Invertebrate fossils are apt to be found along the banks of streams and gullies and in railroad and highway cuts. Many of the limestones bear large numbers of fusulinids or crinoid stems, and the shales may contain many corals, brachiopods, and mollusks. The best collecting will, of course, be found where the rocks have been sufficiently weathered.

Fig. 8. Sketch of typical crinoidal limestone from the Pennsylvanian of north Texas.
Typical invertebrate fossils are foraminifera (principally fusulinids), corals (especially the solitary or “horn” corals), brachiopods, bryozoans (the lacy and branching types are most common), pelecypods, gastropods (exhibiting a variety of coiling), cephalopods (nautiloids and goniatites predominate), and crinoids, which in many areas are found in thick crinoidal limestones (fig. 8). Some typical Pennsylvanian fossils are illustrated in Plates 14, 15, 17, 18, 19, 20, 21, 24, 32, and 35.
Permian rocks are found in widely separated areas in Texas. The best exposed section of marine Permian rocks is found in the Glass Mountains of Brewester County, and many of these rocks are very 42 fossiliferous. The original shell material of some of the Permian fossils of this area has been replaced by siliceous material which is very well preserved. These silicified fossils are removed from the limestone by solution in acid, and some most remarkable specimens have been recovered in this manner (Pl. 3). Brachiopods are the most common fossils, but corals, bryozoans, and mollusks have also been recovered.
Extensive Permian exposures occur also in the central part of the North-Central Plains region. These rocks were formed from sediments of both marine and continental origin and some of them are fossiliferous. The marine rocks contain a variety of invertebrate fossils including brachiopods, pelecypods, gastropods, and ammonoids. Those rocks representing terrestrial deposits contain vertebrate remains at many localities, and numerous amphibians and primitive reptiles (Pl. 40) have been collected from them.
Mesozoic rocks occur over a wide area of Texas and include exposures of Triassic, Jurassic, and Cretaceous age. Many of the Upper and Lower Cretaceous outcrops are quite fossiliferous and easily accessible and thus of considerable interest to many amateur collectors.
Triassic rocks crop out in parts of the High Plains, the Glass Mountains of Trans-Pecos Texas, and parts of Pecos, Crockett, Upton, Reagan, and Glasscock and other west Texas counties. These are predominantly nonmarine rocks consisting of conglomerates, sandstones, shales, and some gypsum beds.
Triassic fossils are almost exclusively vertebrates, although some poorly preserved plant and invertebrate remains have been reported. Fossil vertebrates of the Texas Triassic include phytosaurs (Pl. 42), crocodiles, amphibians, and fish.
In Texas, surface exposures of Jurassic rocks are known only from Malone Mountain in southwestern Hudspeth County. The rocks there are limestones, shales, sandstones, and conglomerates. Fossils reported from that locality include marine and fresh-water pelecypods, fresh-water gastropods, and ammonites.
Rocks of Cretaceous age are widely distributed in Texas and represent one of the more important rock systems of the State. Cretaceous outcrops occur in central Texas, north Texas, the Edwards Plateau, parts of the High Plains, the Gulf Coastal Plain, and Trans-Pecos Texas.
As mentioned earlier, the Texas Cretaceous has been divided into the Lower Cretaceous (Comanche series) and Upper Cretaceous (Gulf series). These rocks consist primarily of marls (a type of calcareous clay), shales, chalks, and limestones, but sands and conglomerates also occur. Cretaceous rocks occur on the surface of about 28 percent of Texas, and many of the larger cities of the State are situated on Cretaceous strata.
Many of the Gulf and Comanche formations contain fossils which are of interest both to amateur and professional paleontologists. Because of their wide distribution in and near large population centers, Cretaceous outcrops can be conveniently visited by many amateur fossil collectors. The fossils are usually abundant and varied, and some are well preserved. Although numerous kinds of fossils may be collected, the more common forms are cephalopods, pelecypods, gastropods, and echinoids. Some of the more typical Cretaceous fossils are shown in Plates 16, 21, 25-28, 32, 33, 35, and 36.
Cretaceous fossils are more commonly found in shales and chalky limestones. Fossiliferous outcrops of these rocks can be found along many streams, roads, and highways of central Texas, north Texas, and the Edwards Plateau. Outcrops which have been weathered are more likely to provide good collecting. In general, collecting is poor in areas covered with heavy vegetation or recent stream deposits. Good 43 collecting localities are outcrops which have a fairly steep slope with a covering of weathered rock material and a minimum of vegetation. One should move slowly from the base of the slope upward while searching the ground for any evidence of fossils, and particular attention should be given to any small gullies since these often contain fossils that have been washed out of upper beds in the exposure.
Cenozoic rocks are widespread in Texas but occur primarily in a broad belt along the Gulf Coastal Plain. In addition, there are exposures of nonmarine Cenozoic strata in the High Plains, North-Central Plains, and Trans-Pecos region. There are also many exposures of Cenozoic igneous rocks in Trans-Pecos Texas.
Rocks of Cenozoic age occur in more than one-third of Texas and consist of conglomerates, sands, clays, and some limestone and lignite beds.
Extensive exposures of Tertiary rocks trend northeast-southwest in a broad band across the Gulf Coastal Plain area. These strata, consisting of sands, clays, and poorly consolidated limestones, are underlain by Cretaceous rocks.
Invertebrate fossils are common in certain Tertiary formations and pelecypods, gastropods, and corals are the predominant forms. In general, however, fossiliferous exposures are of local occurrence and most of the Tertiary formations are unfossiliferous. Those Tertiary invertebrates that are present, however, are often well preserved and represent many interesting types (Pls. 16, 22, 23, 29, 30, 31).
Tertiary invertebrate fossils are commonly found in sands, clays, and marls. Many of these sands and marls have a green color which is due to the presence of glauconite (a green mineral containing iron and closely related to the micas). At certain localities on the Gulf Coastal Plain the glauconite marls and sands of the Weches and Crockett formations contain large numbers of well-preserved clams, snails, and corals. Fossiliferous exposures of Tertiary rocks are sometimes found in road cuts, but better exposures may be found along the banks of rivers and creeks. Certain bluffs along the Brazos, Sabine, and Trinity rivers are well-known Tertiary fossil collecting localities. Many of these better localities are listed in some of the Bureau of Economic Geology bulletins included in the bibliography of this publication (pp. 109-110).
Quaternary deposits of Pleistocene age (geologic time scale, Pl. 1) are found in many parts of Texas and consist of sands, clays, and gravels.
These rocks are distributed along the Gulf Coast in a belt from 50 to 100 miles wide. They occur also as stream terraces in the Edwards Plateau and North-Central Plains regions. In addition, Quaternary sands and gravels are widely distributed over the surface of much of Trans-Pecos Texas. There are also fossiliferous Pleistocene strata in the High Plains region.
Invertebrate fossils are rare in Pleistocene rocks, but some fresh-water and terrestrial mollusks occur. Vertebrate remains, however, are abundant in many localities, and large numbers of horses, camels, mammoths, and other mammals (Pls. 46-49) have been collected. Fossil bones and teeth (figs. 25, 26, p. 104) are commonly found in the gravels and sands of many of the river terraces of the State.
The beginning fossil collector is usually amazed by the many different plants and animals that have left some trace of their existence. In order to understand these different types of prehistoric life, it is necessary to know something about the organisms that are living today.
This handbook discusses the more important groups of plants and animals which have left some sort of paleontological record, and each major group begins with a discussion of the more simple organisms and continues through the more advanced forms. Because scientific workers do not always agree on exactly the same classification, the system adopted in this handbook contains the latest ideas of several workers. It is simple enough to understand, yet complete enough to help one know and classify his fossils. It should be noted that this classification may differ in some respects from that of certain older paleontological publications. Therefore, it has seemed advisable to list other names for some of the groups that are discussed.
In some instances, the brief descriptions and illustrations of each group will enable the collector to make a preliminary identification of his fossils. For more detailed information about each group, the reader should refer to “Books About Fossils” (pp. 108-110).
This part of the handbook begins with a brief summary of the major groups of the plant kingdom, followed by a discussion of the characteristics and relative paleontological importance of the various invertebrate animals. Emphasis is placed on the invertebrates because this type of fossil is most commonly collected by the amateur. Finally, there is a general review of the vertebrates.
Plant fossils are usually fragmental and poorly preserved, and this tends to discourage most amateurs from an active interest in paleobotany. However, in spite of these problems, much is known of the evolution of plants, and plant fossils provide much information about life of the past. In addition, certain plants are of considerable value as indicators of ancient climatic conditions, and their remains have played a large part in the formation of vast coal deposits.
In the following classification only the larger taxonomic groups are discussed. Notice that the term division has been used in place of the term phylum as used in the animal kingdom. This usage is now preferred by many botanists and paleobotanists.
Thallophytes are simple plants without roots, stems, or leaves. They include the fungi, algae, and diatoms (Pl. 12). Diatoms are microscopic fossils that are found in many of the rocks of Texas, and they are quite abundant in Recent sediments as well. Certain of the Paleozoic limestones of central Texas contain banded spherical masses of algae called “algal biscuits.” Although not particularly useful fossils, thallophytes have a long geologic history and are known in rocks ranging from Precambrian to Recent in age.
The bryophytes are simple rootless plants and include the mosses and liverworts. Although more complex, the bryophytes resemble the algae in some respects. They are uncommon fossils, but undoubted bryophytes (liverworts) have been reported from rocks as old as Mississippian.
This division has been divided into four subdivisions, among which are many of the more common living and fossil plants. Such important plants as the ferns, evergreens, hardwood trees, and the flowering plants are all tracheophytes. Among the more common and abundant fossil tracheophytes are the ferns, cycads, and Gingko, in addition to such important “coal plants” as the scale trees, club mosses, and scouring rushes (Pls. 12, 13). The latter commonly occur in many of the world’s great coal deposits, and their remains make up a large part of the coal. Plant fossils of this type may be collected in the dumps around some of the abandoned coal mines in north-central Texas and from other Pennsylvanian rocks in north and Trans-Pecos Texas.
Plate 11
GEOLOGIC RANGE OF THE MAJOR GROUPS OF PLANTS AND ANIMALS
The bands give some indication of the geologic range and relative abundance of the major groups of plants and animals. An increase in the width of the range band corresponds to a relative increase in numbers during the corresponding portion of geologic time.
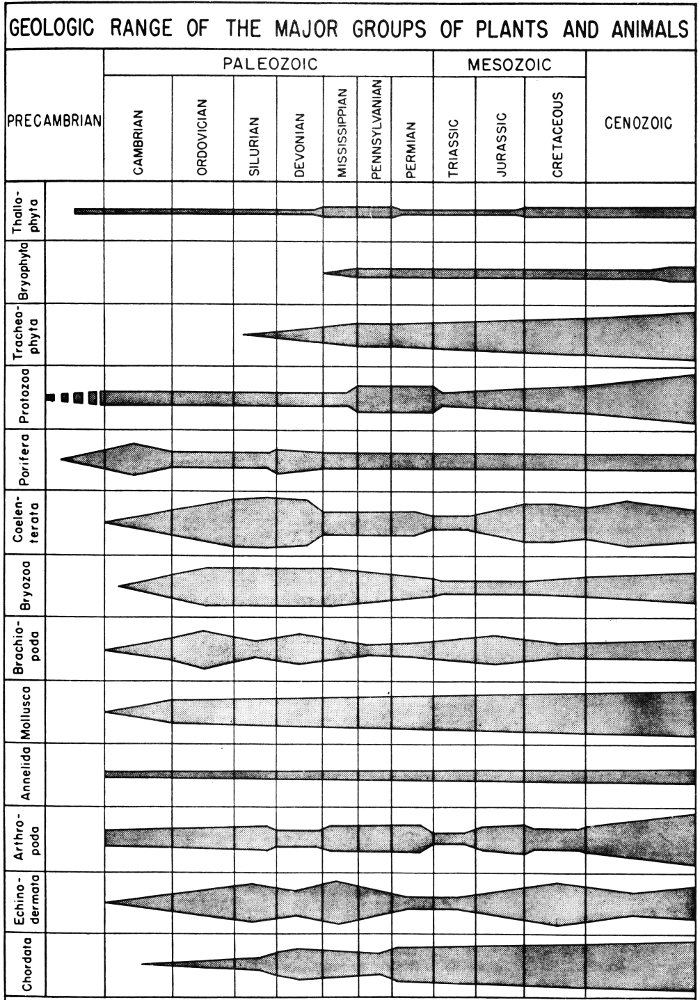
Plate 12
FOSSIL PLANTS

THALLOPHYTES
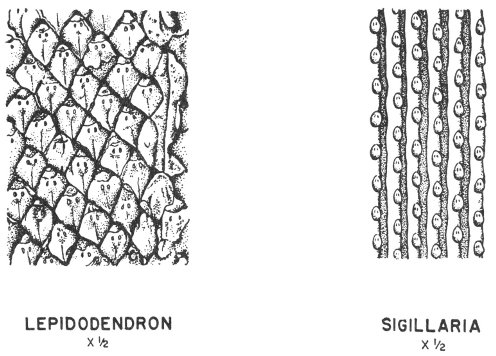
TRACHEOPHYTES
Plate 13
FOSSIL PLANTS
TRACHEOPHYTES

Fairly well-preserved plant remains may also be collected from the Woodbine group of the Upper Cretaceous in north Texas, and fossil wood, most of it silicified, has been reported from rocks of almost all ages and in almost every section of the State. In addition, some of the carbonaceous clays and shales of east Texas contain large assemblages of plant leaves, which in some places are well preserved.
It is also possible to find the fossilized remains of seeds, spores, and pollen. Because of their small size, these minute remains are not destroyed by the drill bit and can be brought out of deep wells without being damaged, and for this reason they are a valuable tool for the micropaleontologist.
The fossilized remains of animals are very common in many of the sedimentary rocks of Texas. These remains are of many different kinds and represent the fossils of such diverse organisms as the shell of a tiny one-celled animal or the bones or tusk of a huge elephant. The fossils most commonly found, however, are the remains of invertebrate animals such as clams, snails, and corals, and it is this type of fossil that attracts the interest of most amateur collectors.
It is not always easy to tell whether certain organisms are plants or animals, and because of this some scientists have suggested that these “in-betweens” be placed in a separate kingdom—the Protista. The protistans are primarily unicellular organisms and are represented by such forms as bacteria, algae, diatoms, and the protozoans (see below). But in this publication, only the plant and animal kingdoms are recognized.
This phylum is composed of simple one-celled animals many of which have no shell or external body covering. Some, however, have external hard parts that can become fossilized, and these forms are quite useful microfossils.
This class contains a group of one-celled animals which may secrete an exoskeleton (external protective covering) of chitin, silica, or calcium carbonate. Included in this class are foraminiferans (commonly called forams) and radiolarians.
Members of this order secrete tiny chambered shells which are very useful microfossils. The forams are predominantly marine organisms and have shells composed of chitin, silica, or calcium carbonate. In addition, some forms construct a shell of sand grains or some other material which is cemented together by a sticky substance that is secreted by the animal.
Forams are very abundant in the rocks of Texas and particularly so in rocks of Mesozoic and Cenozoic age. The most numerous and easily observed Paleozoic foraminiferans are the fusulinids (fig. 9a), and their small spindle-shaped remains are very abundant in many of the Pennsylvanian limestones of north-central and Trans-Pecos Texas. Some typical Texas forams are illustrated in figure 9.
The radiolarians (fig. 10) have delicate spine-covered shells composed of silica, and their remains are very abundant in certain recent marine sediments. They may also be found as fossils and have been reported from Devonian and Permian rocks in 49 Trans-Pecos Texas, and probable radiolarians have been reported from still younger beds.

Fig. 9. Typical Texas Foraminifera (all greatly enlarged). (a) Fusulina (Pennsylvanian). (b) Robulus. (c) Globigerina. (d) Frondicularia. (b-d, Cretaceous).
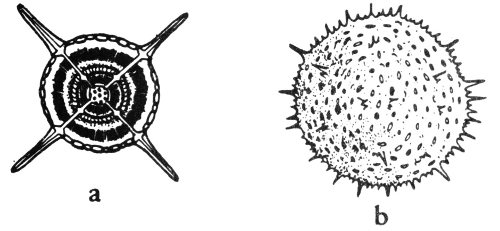
Fig. 10. Typical radiolarians (greatly enlarged). (a) Actinomma (Recent). (b) Porodiscus (Eocene).
These are sponges and are the simplest of the many-celled animals. Living sponges secrete a skeleton which may be composed of chitin, silica, or calcium carbonate. These substances are commonly found in the form of spicules—tiny hard parts that are used to help support the soft tissues of the animal. These spicules take on a variety of shapes (Pl. 14) and are occasionally found as microfossils in some marine sediments.
Although sponges are not particularly common fossils, their remains occur in some parts of the State. Sponges have been collected from Paleozoic and Mesozoic formations of north and Trans-Pecos Texas, and their spicules have been reported from well cuttings.
The coelenterates are multicelled animals which, though more complex than the sponges, are rather primitive animals. The living animal is characterized by a sac-like body cavity, a definite mouth, and tentacles which bear stinging cells. Some forms, for example, the jellyfishes, have an umbrella-shaped body and are single free-moving organisms. Others, like the colonial corals, are composed of many individuals living together in a colony.
Most zoologists and paleontologists recognize three classes of coelenterates: (1) the Hydrozoa, containing the small animals known as hydroids, (2) the Scyphozoa, which includes the jellyfish, and (3) the Anthozoa, which includes the corals and sea anemones. Because of their extreme fragility and lack of hard parts, hydrozoans and scyphozoans are not commonly found as fossils. They do, however, have a long geologic history and may be preserved when unusual conditions of fossilization occur. The anthozoans, especially the corals, are by far the most important class geologically, and these forms have left a very good paleontological record.
This class is composed of a group of exclusively marine organisms and includes the corals and sea anemones. The coral animal, or polyp, secretes a cup-shaped calcareous (limy) exoskeleton. This skeleton, called a corallite, is usually divided by radial partitions called septa. The polyp lives in the calyx, which is the central bowl-shaped depression in the top of the corallite (fig. 11a).
Solitary corals form an individual corallite for each polyp, and because of their shape these may be given such names as “horn corals” (Lophophyllidium, Pl. 15) or “button corals” (Micrabacia, Pl. 16). Colonial or compound corals (Pl. 15) live together in colonies, which are formed of many individual skeletons attached to each other (fig. 11b), and the compound 50 mass of coral skeletons formed in this manner is called a corallum. Fossil corals commonly occur in many marine limestones and in places constitute a large portion of the rock.
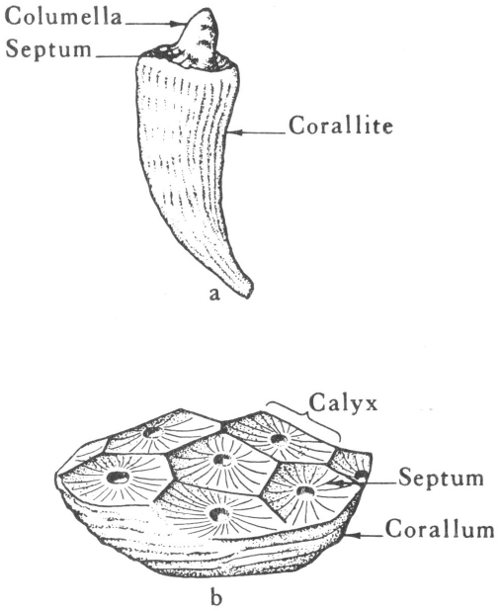
Fig. 11. Morphology and principal parts of corals. (a) Solitary or “horn” coral. (b) Colonial or compound coral.
The class Anthozoa has been divided into several subclasses, but only one, the Zoantharia, is of paleontological importance.
Most corals and all sea anemones belong to this subclass. Zoantharians are either colonial or solitary and, because most of them possess a hard preservable exoskeleton, they are the most important group of anthozoans geologically. The various orders of the subclass Zoantharia are discussed below.
These are corals in which the septa are arranged in cycles of four. Both solitary and colonial forms occur, and they are found only in rocks of Paleozoic age. Rugose corals are abundant in many of the Paleozoic formations of Texas, and two of the more typical forms (Lophophyllidium and Caninia) are illustrated in Plate 15. Members of this order have been placed in the subclass Tetracoralla of older classifications.
The scleractinians are solitary or colonial corals in which the septa grow in multiples of six, and they are the most important and abundant of the modern corals. These corals were the dominant reef builders of Mesozoic and Cenozoic seas, and their remains are common in many of the marine formations of the State. Plate 16 illustrates some typical scleractinian corals from the rocks of Texas. This order has also been referred to as subclass Hexacoralla, and its members have been called hexacorals.
These are corals that are now extinct but are known from fossils in both Paleozoic and Mesozoic rocks. Tabulate corals are characterized by horizontal partitions called tabulae, and septa are absent or poorly developed. The tabulates were the most abundant reef-building corals during Paleozoic time and are well known as fossils. Because of certain similarities with other anthozoans, some paleontologists have treated the Tabulata as a distinct subclass rather than as an order of the Zoantharia.
Tabulate corals are not uncommon in many of the Paleozoic rocks of Texas, and two of these (Cladochonus and Striatopora) are illustrated in Plate 15.
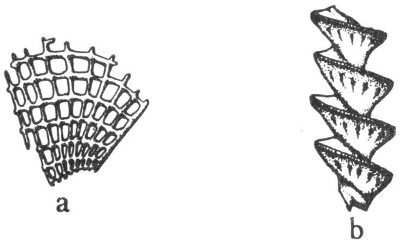
Fig. 12. Two types of bryozoans or “moss animals.” (a) Section of the lacy type bryozoan. (b) The spiral axis of Archimedes (Mississippian).
Bryozoans are colonial animals that are often referred to as “sea mats.” They have been called this because they are commonly found matted on shells, rocks, fossils, and other objects. The living animal is quite small, has a tentacle-bearing ridge surrounding the mouth, and secretes a tiny cup-like exoskeleton composed of calcareous or chitinous material. These little chambers, known as zooecia (or autopores), are seen as small pits on the surface of the bryozoan colony (Rhombopora, Pl. 17). The zooecia grow together to form the bryozoan colony, and some fossil colonies grow to be as much as 2 feet across. Such colonies may be spiral (fig. 12b), branching, or lace-like (fig. 12a), and the latter two types are very common in many of the fossiliferous strata of Texas. Undoubted bryozoan fossils have been recorded in rocks of Lower Ordovician age, but questionable Cambrian forms have also been reported. Bryozoans are abundant in the seas of today, but only a few forms inhabit fresh waters.
Plate 14

SPONGE SPICULES
(GREATLY ENLARGED)
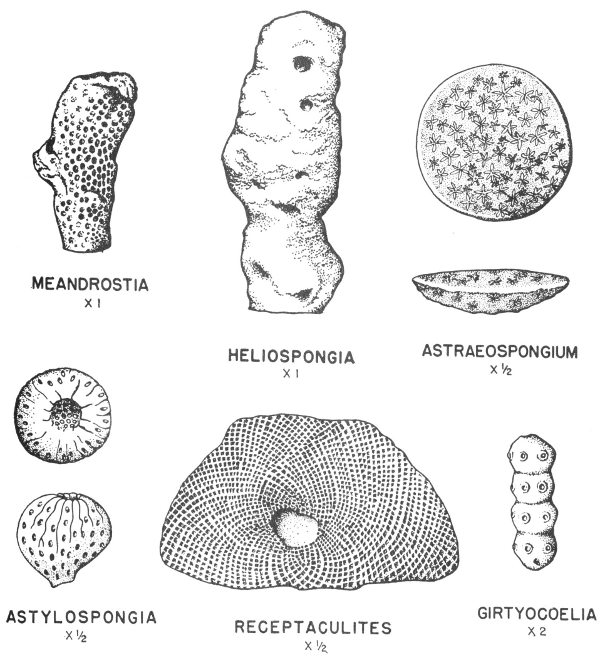
PALEOZOIC SPONGES
Plate 15
PENNSYLVANIAN CORALS
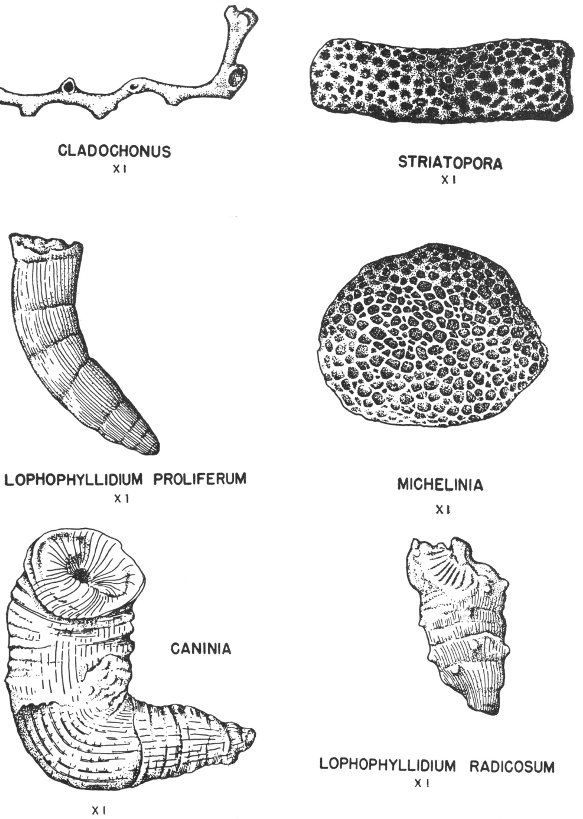
Plate 16
CRETACEOUS CORALS
TERTIARY CORALS
In Texas one may expect to find bryozoan remains in the Pennsylvanian rocks of north-central and Trans-Pecos Texas where they are abundant in certain of the marine shales and limestones. Bryozoans may also be collected from some Cretaceous and Tertiary beds, but their remains are small and fragmental and they are easily overlooked. Bryozoans have also been found matted on the shells of fossil mollusks and other invertebrates.
The brachiopods are a large group of exclusively marine organisms with shells composed of two pieces called valves (fig. 13). These valves are usually composed of calcareous or phosphatic material and enclose and protect the soft parts of the brachiopod animal. The soft parts are composed of muscles, the mantle (which secretes the valves), digestive, respiration, reproductive, and excretory organs, and the tentacle-bearing lophophore.
In adult life the brachiopod is attached to the sea bottom by means of a fleshy stalk called the pedicle (fig. 14), and this is usually extruded through a hole (the pedicle foramen) which is located in the ventral or pedicle valve. The upturned area which is usually present on the pedicle valve is called the beak. The other valve, known as the dorsal or brachial valve, is usually the smaller of the two (fig. 13b). The two valves are opened by means of muscles, and since death results in relaxation of these muscles, fossil brachiopods are typically found with valves closed.
Brachiopods vary greatly in size and shape and exhibit a wide variety of ornamentation, such as spines, ribs, nodes, and other structures. They are abundant fossils in many of the Paleozoic rocks of Texas but are relatively rare in Mesozoic and Cenozoic formations.
The phylum has been divided into two subclasses, the Inarticulata and the Articulata. This classification is based upon the nature of the hinge-line—the edge of the shell where the two valves articulate.
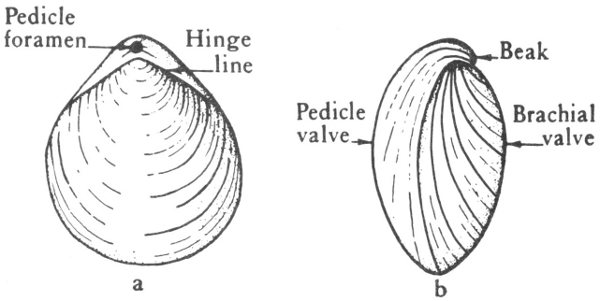
Fig. 13. Morphology and principal parts of articulate brachiopods.
The members of this class are rather primitive and have a long geologic history. These brachiopods have valves which are not provided with hinge teeth, the valves being held together by muscles, and a hinge-line is lacking (fig. 14). Most inarticulate brachiopods are circular or tongue-like in shape and commonly composed of chitinous and phosphatic material. Inarticulate brachiopods range from Lower Cambrian to Recent in age but were never as common as the articulate brachiopods, which are described below. Brachiopods belonging to this class have been recorded from several Paleozoic formations in Texas (Pl. 17, Lingula, Apsotreta, Angulotreta).
Plate 17

PENNSYLVANIAN BRYOZOANS
CAMBRIAN BRACHIOPODS
MISSISSIPPIAN BRACHIOPODS
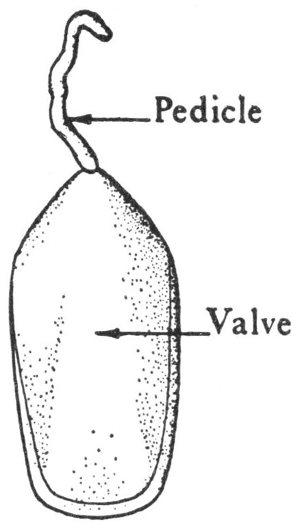
Fig. 14. Lingula, a typical Recent inarticulate brachiopod showing extended pedicle.
Articulate brachiopods have a well-defined hinge-line (fig. 13a). One valve has well-developed teeth which articulate with sockets in the opposing valve, and there is a well-developed muscle system which aids in opening and closing the shell. Articulate brachiopods are characterized by calcareous shells which are typically of unequal size and a wide variety of shapes (Pls. 18, 19). The class has been divided into several orders which have been established primarily on the nature of the pedicle foramen and the nature of shell growth.
Articulate brachiopods range from Lower Cambrian to Recent in age and are particularly abundant in certain Pennsylvanian formations of north-central and Trans-Pecos Texas. They are also present in certain other fossiliferous strata of Paleozoic age but are less abundant and not as well preserved. The only Cretaceous brachiopod that is found in large numbers is Kingena wacoensis (Roemer) (fig. 15), which is particularly abundant in certain formations in the upper part of the Comanche series.
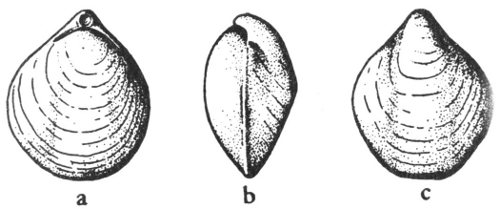
Fig. 15. Kingena wacoensis, a common Cretaceous brachiopod. (a) Dorsal view. (b) Lateral view. (c) Ventral view.
The phylum Mollusca encompasses a large group of aquatic (water-dwelling) and terrestrial (land-dwelling) invertebrates which includes such familiar forms as the snails, clams, oysters, squids, and octopuses. Most mollusks possess a calcareous shell that serves as an exoskeleton, and these hard parts are well adapted for preservation as fossils. However, some mollusks (the slugs) have no shells, and others (the squids) have an internal calcareous shell. Because of their relative abundance and great variety, mollusks are particularly useful fossils. Moreover, the remains of certain mollusks, such as the oysters, are important rock builders.
The phylum Mollusca has been divided into five classes:
1. Amphineura—the chitons or sea-mice; shell composed of eight valves or plates; not a common fossil. Ordovician to Recent.
2. Scaphopoda—the tusk-shells; shell composed of a single tusk-like valve; generally not a common fossil but locally abundant in certain Cenozoic formations. Devonian to Recent.
3. Gastropoda—the snails and slugs; slugs are without shells, snails have a single-valved shell which is typically coiled; common fossils in Paleozoic, Mesozoic, and Cenozoic rocks. Cambrian to Recent.
4. Pelecypoda—clams, mussels, oysters, scallops; shells composed of two valves, usually, but not always, of equal size; common fossils, especially in Mesozoic and Cenozoic rocks. Cambrian to Recent.
5. Cephalopoda—squids, octopuses, the pearly nautilus, and the ammonoids (extinct); shell of one valve, usually coiled and partitioned by septa; valuable fossils, especially in Paleozoic and Mesozoic rocks. ?Cambrian, Ordovician to Recent.
Plate 18
PENNSYLVANIAN BRACHIOPODS
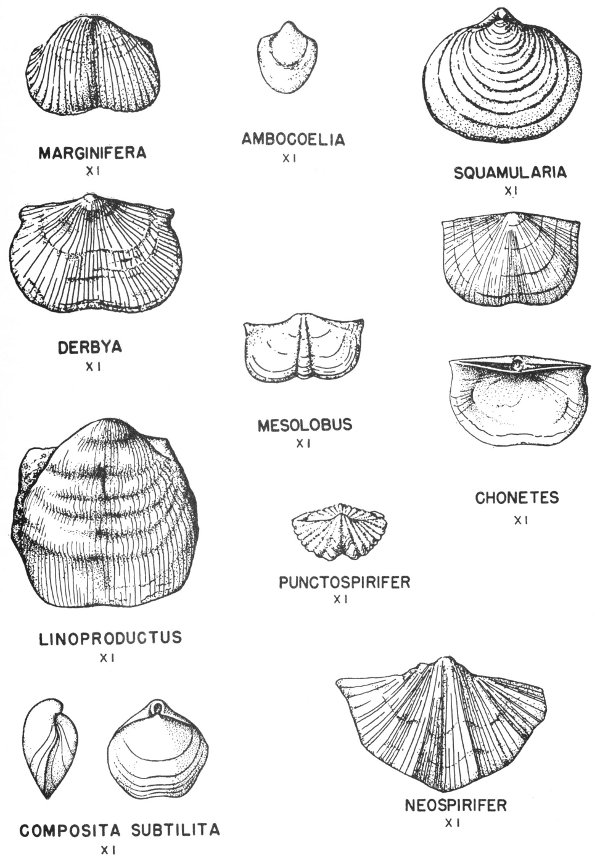
Plate 19
PENNSYLVANIAN BRACHIOPODS
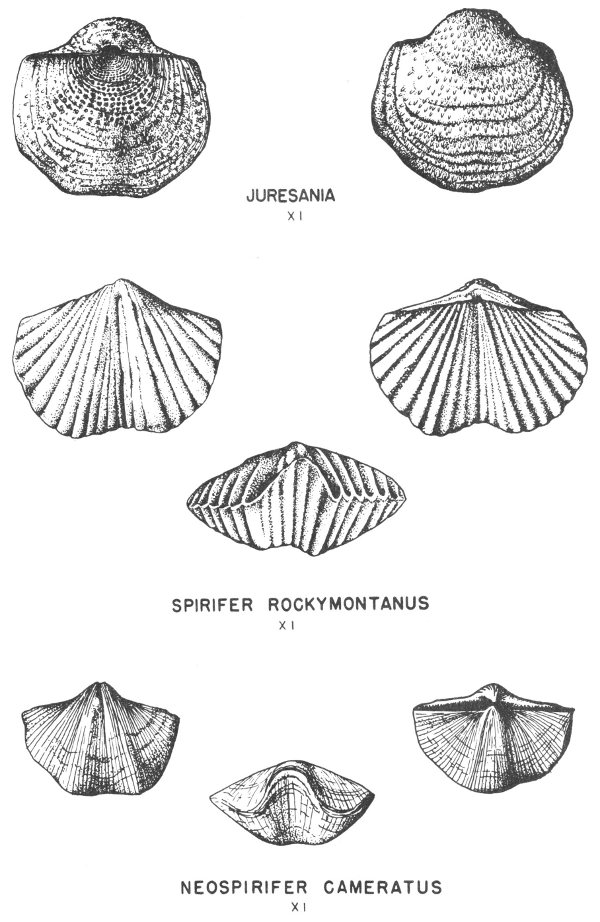
Of these five classes, only the Gastropoda, Pelecypoda, and Cephalopoda are discussed herein.
The typical gastropod has a spirally coiled, single-valved, unchambered shell. This shell encloses a soft body possessing a well-defined head with a pair of eyes and one or two pairs of tentacles. Most gastropods have gills and live in shallow marine waters, but some inhabit fresh water. Others are land-dwelling forms and breathe by means of lungs.
Gastropod shells, both Recent and fossil, exhibit a great variety of size, shape, and ornamentation. Such shells may be cone-shaped, spirally coiled, flat, turreted, or cylindrical. The shell is commonly wound in a spiral around a central axial pillar (the columella). The closed pointed end of the shell is called the apex, and each turn of the shell is called a whorl (fig. 16). The last-formed and largest whorl is called the body whorl, and this whorl contains the aperture—the opening of the shell. The combined whorls exclusive of the body whorl are known as the spire. The inner and outer margins of the aperture are designated the inner lip and the outer lip, respectively. In some snails the aperture is closed by means of the operculum—a calcareous or horny plate attached to the foot of the animal. This plate effectively seals the aperture when the animal is withdrawn into its shell. Some gastropods have shells that are loosely coiled, and in these forms the columella is absent. If the whorls of such shells are not in contact on the inner surface, this leaves an open space which is called the umbilicus (fig. 16a). The umbilicus is commonly seen as an opening in the base of the gastropod shell, but in some forms the umbilical opening may be partially or completely covered by a thick growth of shell called the callus.
Many gastropods, particularly those of the Texas Cretaceous, are commonly preserved as internal or external molds. This type of preservation occurs after the death of the animal, and the decomposition of the soft parts enables the shell to become filled with sediment. This filling later becomes solidified, and the outer shell may eventually be removed by weathering or solution. This type of internal mold is called a steinkern and normally does not reflect any external shell characteristics (Pl. 2). In some of the Pennsylvanian and Tertiary formations, however, gastropods may be collected with the original shell in an excellent state of preservation.
Plates 20-23 illustrate some typical Paleozoic, Mesozoic, and Cenozoic gastropods.
The pelecypods possess a shell composed of two calcareous valves (fig. 17) which enclose the soft parts of the animal. Members of this class live exclusively in an aquatic habitat and are most abundant in marine environments. Most pelecypods are slow-moving bottom-dwelling forms, but some, like the oysters, are attached. Still others, for example, the scallop or Pecten, are swimmers. The Pelecypoda include such familiar saltwater forms as the clams and oysters, as well as the common fresh-water mussel. Pelecypods range from Cambrian to Recent in age but are more abundant in Mesozoic and Cenozoic rocks.
The living animal is aquatic, with well-developed soft parts and a muscular, commonly hatchet-shaped foot. The soft mantle encloses the body and secretes the shell, and in some pelecypods part of the mantle is developed into the incurrent and excurrent siphons. The incurrent siphons bring fresh water and food into the mantle cavity, and waste products are passed out through the excurrent siphons. Respiration is by means of gills within the mantle cavity.
The typical pelecypod valves are of equal size and form, but some, such as the scallops and oysters, have two valves of unequal size and shape. The valves are hinged and held together by a tough elastic ligament which runs along the dorsal (top) side of the shell. In addition to the ligament, 60 most forms have teeth and sockets which are located along the hinge-line. The teeth in one valve articulate with the sockets in the opposite valve, and this arrangement gives strength to the hinge.

Fig. 16. Morphology and principal parts of gastropod shells. (a) Low-spired form with umbilicus. (b) Section of spirally coiled shell showing columella.
Most of the pelecypod shell is of calcium carbonate, but the outer layer, or periostracum, of each valve is composed of horny material. The inner surface of the shell is lined with a calcareous layer of pearly or porcelaneous material.
Plate 20
PENNSYLVANIAN GASTROPODS

Plate 21
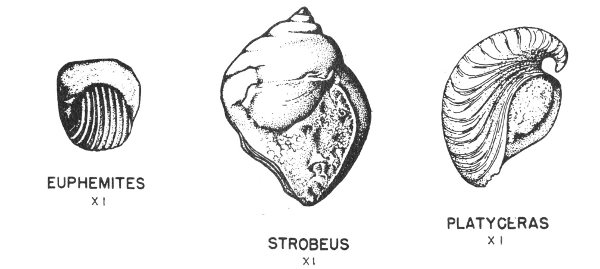
PENNSYLVANIAN GASTROPODS
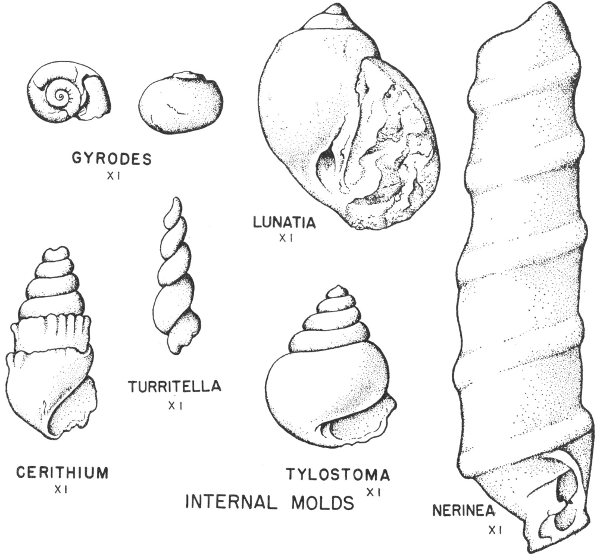
CRETACEOUS GASTROPODS
INTERNAL MOLDS
Plate 22
TERTIARY GASTROPODS
Plate 23
TERTIARY GASTROPODS
The outline of the shell may vary greatly, but most pelecypods are typically clam-like. However, certain forms are round, others are long and narrow, and some have wing-like structures. Most pelecypods have a beak which represents the oldest part of the shell. The beak is commonly located on the anterior (front) end of the shell, and the end of the shell opposite this is designated posterior (the rear). The hinge and ligament are located dorsally (along the top), and the lower margin of the shell where the valves open is called ventral (fig. 17a).

Fig. 17. Morphology and principal parts of a typical pelecypod shell. (a) Exterior view. (b) Interior view.
The inner surface of the shell has certain markings which, along with the shell form and dentition (the nature and arrangement of the teeth and sockets), are important in classification. Muscle scars are present on the inside of most valves; the anterior muscle scars are located near the front of the shell, and the posterior muscle scars are situated near the rear of the shell. These scars mark the place of attachment of muscles which were used to close the shell and aid in locomotion. Along the ventral margin of some shells there is a line or scar which extends from the anterior muscle scar to the posterior muscle scar. This is known as the mantle line or pallial line and marks the place of attachment of the mantle—a soft membranous layer that enclosed the body of the animal. In some pelecypods the dorsal margin of one valve bears a series of hinge teeth which articulate with a similar set of sockets on the other valve (fig. 17b). In addition to hinge teeth, certain species have cardinal teeth which are located below and in front of the hinge teeth.
The exterior of most shells is marked by a series of concentric growth lines (fig. 17a) which mark points of periodic addition of shell material. The external surface of many shells is also marked by various types of ornamentation, such as ribs, nodes, spines, and grooves.
Fossil collectors commonly find only one valve of the pelecypod shell. This is because the shell normally opens when the animal dies, and the valves may easily become separated. Fossil pelecypods are also commonly preserved as external and internal molds, and these are found in fossiliferous strata of almost all ages. Some pelecypods of Pennsylvanian, Mesozoic, and Cenozoic age are found with original shell material that appears to have undergone very little change. Fossil pelecypods are abundant and varied in Texas and are found in most of the fossiliferous formations of the Pennsylvanian, Cretaceous, and Tertiary systems (Pls. 24-31).
These are marine mollusks with or without chambered or solid shells which may be internal or external. The living animal possesses a well-developed head with eyes, horny jaws, and many tentacles fused with the foot. Cephalopods are the most advanced of all mollusks and include the squid, octopus, pearly nautilus, and the extinct ammonoids. Members of this class range from Cambrian to Recent in age but were much more abundant in ancient seas than they are today. Their remains constitute a very useful group of fossils, particularly in Paleozoic and Mesozoic rocks.
Most paleontologists have divided the Cephalopoda into three subclasses, the Nautiloidea, Ammonoidea, and the Coleoidea (known also as subclass Dibranchiata and subclass Decapoda); each of these is discussed below.
The nautiloids are cephalopods with external chambered shells in which the septa (dividing partitions) are simple and have smooth edges. This subclass is represented by a single living genus, Nautilus, and a large number of fossil forms.
In the living Nautilus the shell is composed of calcium carbonate and is coiled in a flat spiral (fig. 18). The interior of the shell is divided into a series of chambers by calcareous partitions called septa. The point where each septum joins the inner surface of the shell is known as the suture. These suture lines (fig. 19a) are not visible from the outside unless the outer shell has been removed, but they are visible on the internal molds of many fossil cephalopods and are of great importance in nautiloid and ammonoid classification. Nautiloids have very simple smoothly curved suture patterns, but ammonoids are characterized by more complex and wrinkled sutures (fig. 19d).
Although the shell of the only type of living nautiloid is coiled, many of the early forms had straight cone-shaped shells (Orthoceras, Pl. 32), and these are common in some of the Pennsylvanian formations of Texas. Fossil coiled nautiloids may be collected in certain of the Cretaceous and Tertiary strata of the State, but their remains are not common. Cymatoceras (Pl. 32) is a coiled fossil nautiloid from the Cretaceous of north Texas.
Plate 24
PENNSYLVANIAN PELECYPODS
Plate 25
CRETACEOUS PELECYPODS
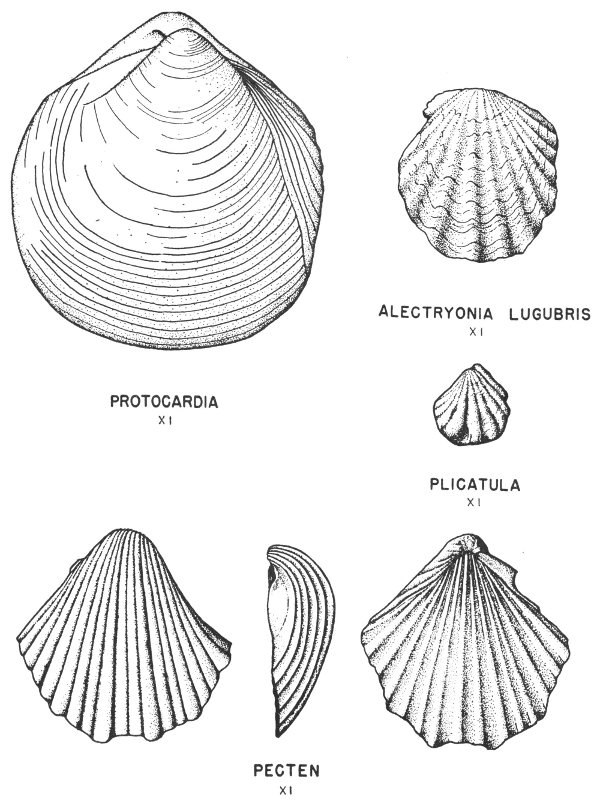
Plate 26
CRETACEOUS PELECYPODS
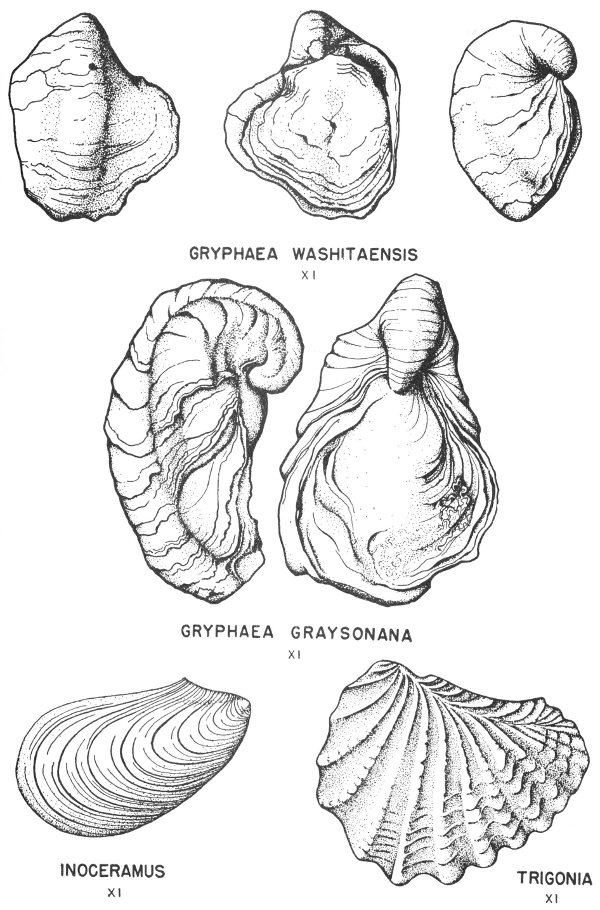
Plate 27
CRETACEOUS PELECYPODS
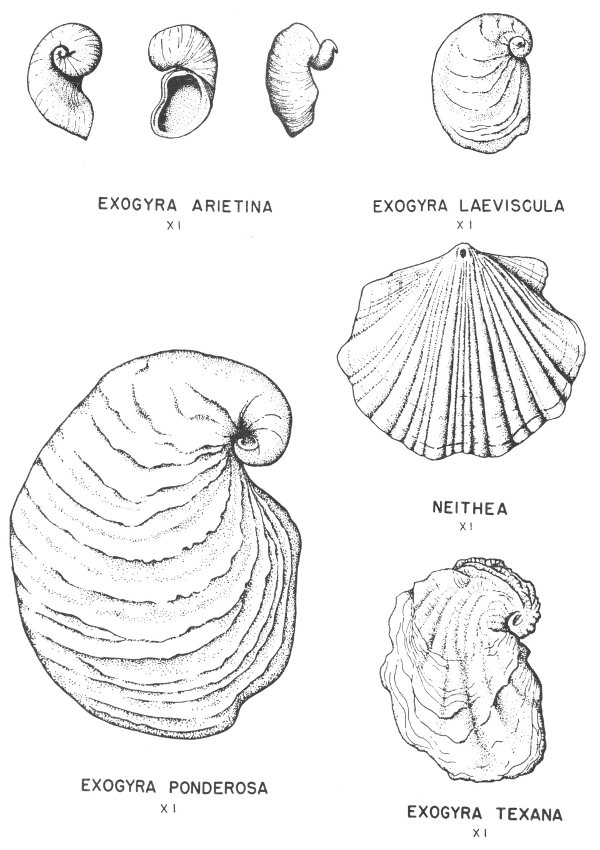
Plate 28
CRETACEOUS PELECYPODS
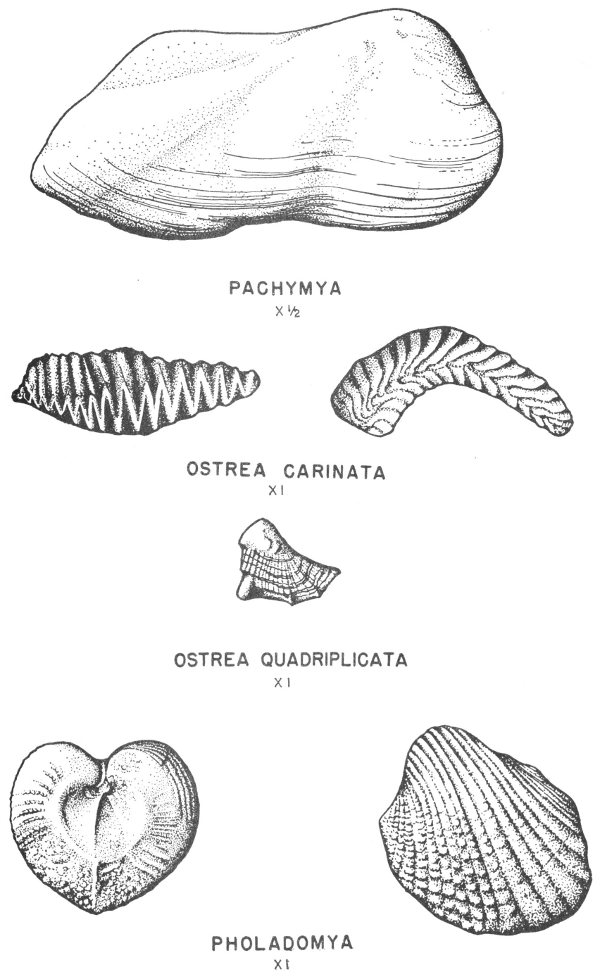
Plate 29
TERTIARY PELECYPODS
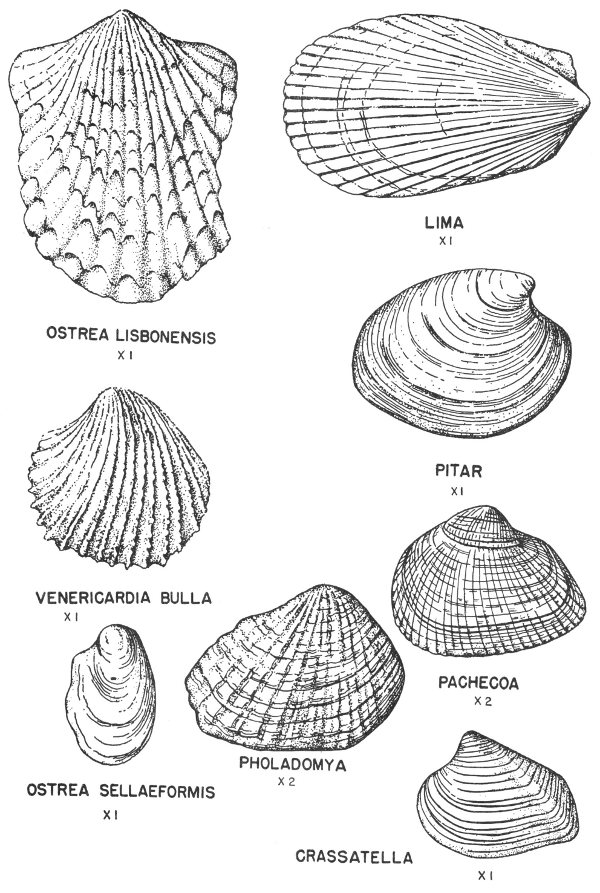
Plate 30
TERTIARY PELECYPODS
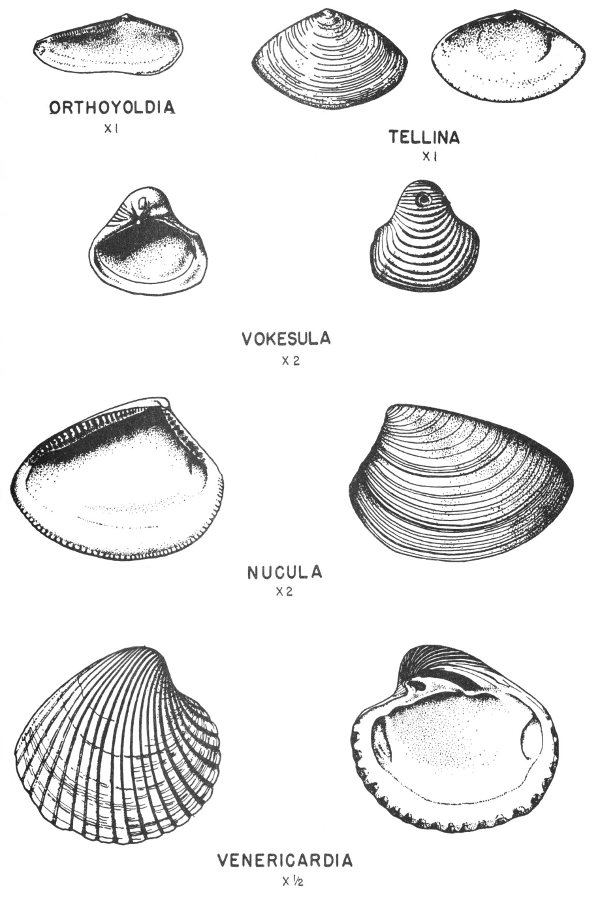
Plate 31
TERTIARY PELECYPODS
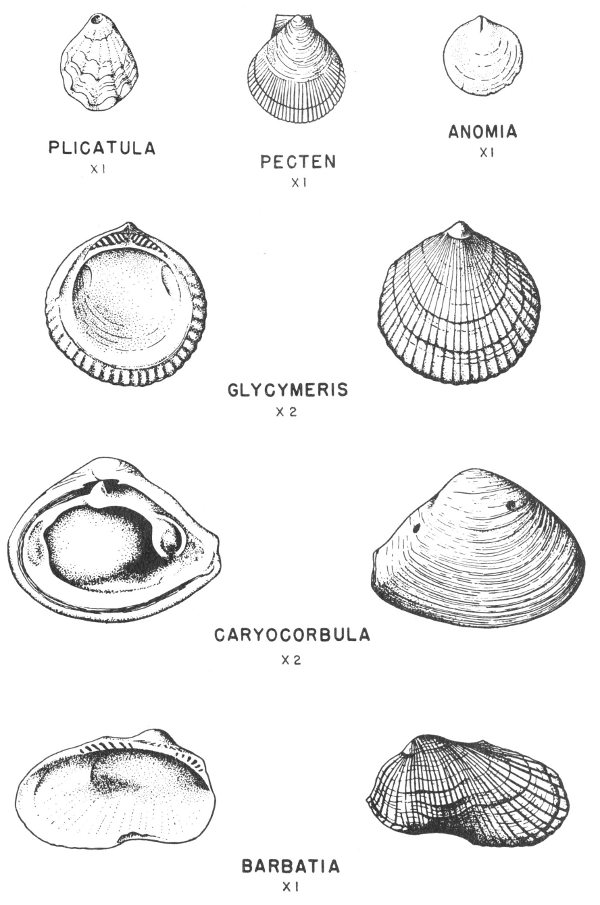
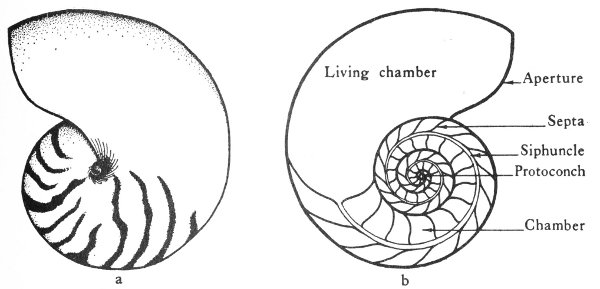
Fig. 18. Morphology and principal parts of the pearly nautilus. (a) Exterior view of a Recent shell. (b) Sectioned view of the same shell to show internal structures.
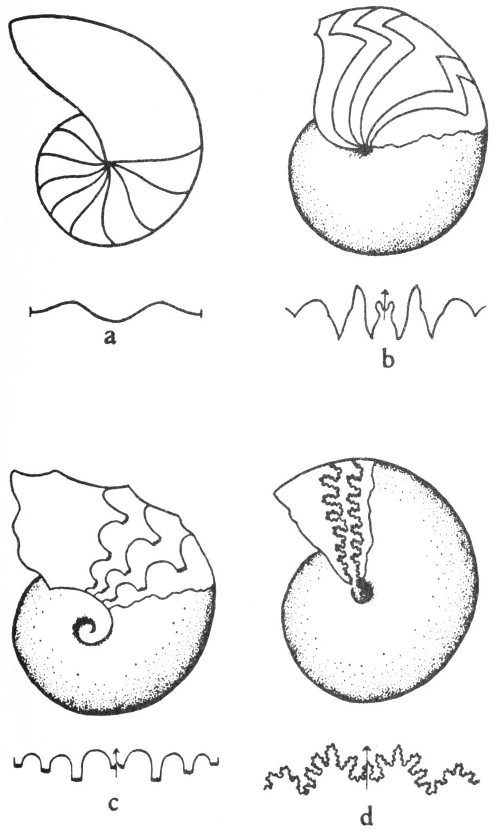
Fig. 19. Characteristic features of the various types of cephalopod sutures. (a) Nautiloid type. (b) Goniatite type. (c) Ceratite type. (d) Ammonite type.
The ammonoids are a group of extinct cephalopods which are related to the nautiloids but are characterized by more complex suture patterns. Members of this subclass have an external partitioned shell which is straight, curved, or spirally coiled (Pl. 33). This group of cephalopods first appeared in Devonian time, became extremely abundant and varied during the Mesozoic, and was extinct by the end of the Cretaceous period.
Most Paleozoic ammonoids are characterized by a combination curved and angular suture pattern, and this type of suture pattern is referred to as goniatitic (fig. 19b). Sutures that are curved and crenulated (marked in places by a series of tooth-like indentations) are known as ceratitic (fig. 19c). Ceratitic sutures first appeared in the Mississippian, became increasingly abundant during the Triassic but were much less abundant during the Cretaceous. The ammonitic suture has a very complexly subdivided pattern (fig. 19d). Cephalopods with ammonitic sutures range from Pennsylvanian to Cretaceous in age and were the most abundant cephalopods of the Mesozoic.
Plate 32
PENNSYLVANIAN CEPHALOPODS
CRETACEOUS CEPHALOPODS
Plate 33
CRETACEOUS CEPHALOPODS
Ammonoids are locally abundant in many of the fossiliferous rocks of Texas and are among the more useful Mesozoic guide fossils. Goniatites may be found in the Pennsylvanian of north-central and Trans-Pecos Texas, and ammonoids with the ceratitic suture pattern can be collected from the Lower Cretaceous of many parts of the State. Cephalopods exhibiting the typical ammonitic suture pattern are abundant in many of the Cretaceous rocks of Texas, and these fossils have contributed much toward an understanding of the Cretaceous strata of this State.
These are squid-like cephalopods characterized by an internal shell or no shell at all. Included in this group are the squids, cuttlefish, octopuses, and the extinct belemnoids, but of these only the belemnoids are useful fossils. Members of this subclass range from Mississippian to Recent in age.
The belemnoids appear to be the oldest and most primitive of the coleoid cephalopods. Their earliest known occurrence is in rocks of Mississippian age, and they were particularly abundant during the Mesozoic. They became extinct at the end of Cretaceous time but have left considerable evidence of their existence in the Mesozoic strata of many parts of the world. Certain forms, because of their abundance and relatively short geologic range, are excellent guide fossils. Belemnoids have been found in the Upper Cretaceous of Texas (Pl. 33) but in general are rare or unknown in most Texas formations.
Members of the phylum Annelida include the segmented worms such as the common earthworm. Annelids are marine, fresh water, or terrestrial and have apparently been common through much of geologic time. Because of their lack of hard parts, most of these worms have left little direct fossil evidence of their activities in the geologic past. Some annelids secrete straight or coiled calcareous tubes, and fossil worm tubes of this sort (fig. 20) are commonly found attached to brachiopods, mollusks, and other objects. Tubes of this nature have been reported from Paleozoic, Mesozoic, and Cenozoic rocks in Texas.
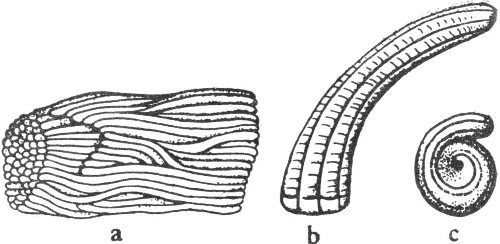
Fig. 20. Types of typical annelid worms. (a) Serpula (×1) (b) Hamulus (×2). (c) Spirobis (×15).
Some annelids have small chitinous jaws and teeth which also may be preserved as fossils. These dental structures are called scolecodonts and are microfossils.
The arthropods are one of the more advanced groups of invertebrates, and they are known from the Cambrian to the Recent (Pl. 34). Modern representatives of this group include the crabs, shrimp, crayfish, insects, and spiders. Arthropods vary greatly in size and shape and are among the most abundant of all animals. They have become successfully adapted to a wide variety of environments and live on land, in water, and in the air. The typical arthropod has a segmented body which is usually covered by a chitinous exoskeleton which, in some forms, contains additions of calcium carbonate. They are highly specialized and well-developed animals in which locomotion is by means of paired jointed appendages.
Although the arthropods are of great importance in nature today, only a few groups are of importance to the paleontologist. Only two of these, the trilobites and the ostracodes, are discussed herein.
The members of this subphylum are extinct arthropods which were most abundant during early Paleozoic time.
The trilobites are a group of exclusively marine arthropods which derive their name from the typical three-lobed appearance of their bodies (fig. 21a). The trilobite body is divided into a central or axial lobe and two lateral lobes. The body of the animal was encased in a chitinous exoskeleton. The top part of this exterior covering, the carapace, is very thick, and it is this part of the trilobite that is usually preserved.
Plate 34
FOSSIL ARTHROPODS
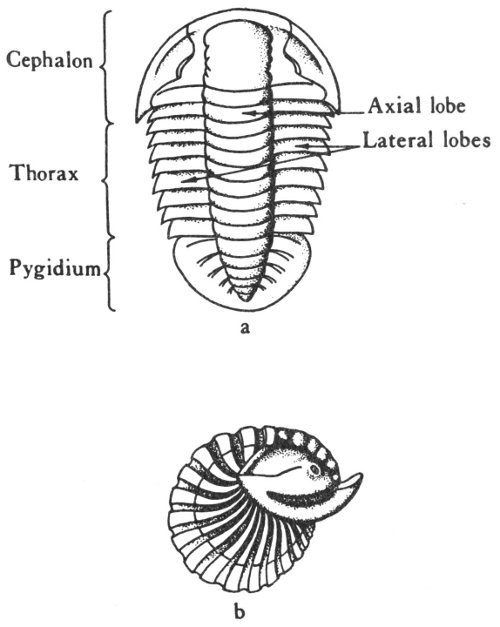
Fig. 21. Morphology and principal parts of trilobites.
The body is also divided into three parts from front to back. Beginning at the front of the animal these divisions are the cephalon or head, the thorax or abdomen, and the pygidium or tail (fig. 21a). The body segments of the thorax were arranged in such a manner as to permit the animal to roll up into a ball, and many trilobites are found in this position (fig. 21b).
Trilobites first appeared in the Cambrian and were extinct by the end of the Permian. They occur sparingly in certain of the Paleozoic rocks of Texas and when found are likely to be fragmental and in a poor state of preservation.
The crustaceans are the crabs, shrimp, crayfish, and ostracodes. Although not abundant, fossil crabs have been described from certain Cretaceous and Tertiary formations of the State (Notopocorystes, Pl. 34). However, the most useful and abundant crustacean fossils are the members of the class Ostracoda.
The ostracodes are small, bivalved, aquatic crustaceans which have the external appearance of small clams (Pl. 34). The remains of these tiny animals are so small that they are best studied under a low-power microscope, and because of their small size they are particularly useful to the micropaleontologist.
Fossil ostracodes range from Ordovician to Recent in age and have been recorded in the Paleozoic, Mesozoic, and Cenozoic rocks of Texas. Their remains are particularly abundant in certain of the Cretaceous and Tertiary marine formations of the State.
The echinoderms are a large group of exclusively marine animals, most of which exhibit a marked five-fold radial symmetry (Pls. 35, 36). Living echinoderms have well-developed nervous and digestive systems, a distinct body cavity, and are a relatively complex group of organisms.
The typical echinoderm has a skeleton composed of numerous calcareous plates which are intricately fitted together and covered by a leathery outer skin (the integument). The echinoderm body often exhibits a typical star-shaped form, but some types may be heart-shaped, biscuit-shaped, or cucumber-shaped.
Members of this phylum range from Cambrian to Recent in age and are abundant as fossils in many of the marine formations of Texas.
The phylum Echinodermata has been divided into two subphyla, the Pelmatozoa (those forms that were attached to sea floor by a stem or a stalk) and the 81 Eleutherozoa (the stemless unattached echinoderms).
These are echinoderms which are more or less permanently attached to the bottom of the sea by means of a stalk which is composed of slightly movable, calcareous, disk-like segments (fig. 23).
Pelmatozoans range from Cambrian to Recent in age, and their fossilized remains are particularly abundant in Paleozoic rocks. The subphylum has been divided into several classes, but only three of these, the Cystoidea, Blastoidea, and Crinoidea, are discussed here. With the exception of the Crinoidea, all of the attached echinoderms are extinct.
These are primitive attached echinoderms which were relatively common in early Paleozoic time. The typical cystoid has a somewhat globular or sac-like calyx (the main body skeleton) composed of numerous, irregularly arranged, calcareous plates (fig. 22b). The plates composing the calyx are usually perforated by pores or slits which were probably used in excretion or respiration. The calyx was attached to the sea bottom by a short stem.
Cystoids range from Cambrian to Devonian in age and were especially abundant during Ordovician and Silurian time. Their remains are rare or absent in the rocks of Texas.
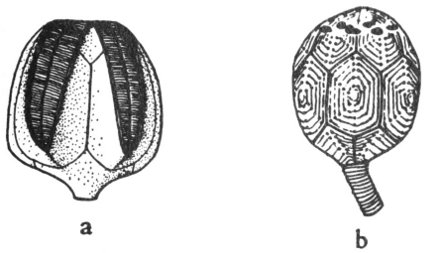
Fig. 22. Two extinct attached echinoderms. (a) Pentremites (Mississippian). (b) Caryocrinites (Silurian).
The blastoids are extinct short-stemmed echinoderms with a small, symmetrical, bud-like calyx. The blastoid calyx is composed of 13 calcareous plates arranged in a typical five-sided pattern (fig. 22a). The mouth is located in the center of the calyx and is surrounded by five openings called spiracles. Five distinct ambulacral or food grooves radiate outward from the mouth.
Blastoids range from Ordovician to Permian in age and were especially abundant during the Mississippian period. No blastoids have been reported from any of the rocks of Texas.
The crinoids are commonly called sea-lilies because of their flower-like appearance. The calyx is composed of symmetrically arranged calcareous plates, and most crinoids have a long stem. Other crinoids are free-swimming in the adult stage and are attached only during the earlier phases of their development.
The crinoid calyx is typically cup-shaped (fig. 23) and five grooves radiate out from its center. These grooves continue outward along the complexly segmented arms and are used as channels to convey food to the mouth.
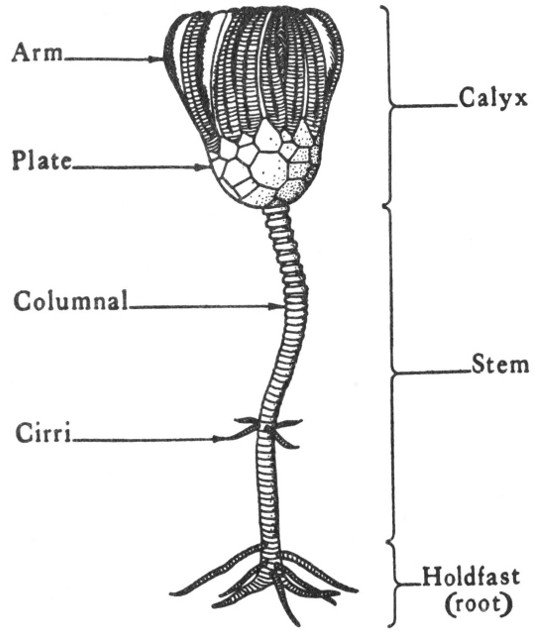
Fig. 23. Typical modern crinoid, or “sea lily,” showing principal parts.
The crinoid stem is attached to the base 82 of the calyx and serves for purposes of support and attachment. This stem consists of a relatively long flexible stalk composed of numerous calcareous disk-shaped segments called columnals (fig. 23; Pl. 35), each of which contains a round or star-shaped opening in its center. Many crinoids have very long stalks (some are as much as 50 feet in length), and when the animal dies the columnals become separated and are scattered about on the ocean floor. Many Paleozoic limestones contain such great numbers of crinoid columnals that they are referred to as crinoidal limestones (fig. 8). Crinoidal limestones occur in some of the Mississippian and Pennsylvanian formations of central Texas and in the Pennsylvanian of north-central and Trans-Pecos Texas.
The stalk is attached to the sea floor or some other object by means of a root system called the holdfast (fig. 23). This structure commonly branches out into the surrounding sediments, and in this manner the crinoid animal is firmly anchored to the bottom of the sea.
Crinoids, like most echinoderms, are gregarious animals—that is, they commonly live together in large numbers, and for this reason great numbers of crinoid remains are commonly found concentrated in relatively small local areas. Most fossil crinoids are found as stem fragments because the more fragile calyx and root system are less likely to be preserved.
The earliest known crinoids have been found in rocks of Ordovician age, and their remains are particularly abundant in Paleozoic rocks. Crinoids are living today but most of them are stemless free-swimming forms called “feather stars,” much less abundant than their Paleozoic ancestors.
The eleutherozoans are free-swimming, bottom-dwelling, echinoderms which have been divided into two classes. The class Asterozoa (star-shaped echinoderms) contains the subclasses Asteroidea (the starfishes) and the Ophiuroidea (the brittle stars). Although they are known as fossils, neither of these groups is of paleontological importance. The class Echinozoa (echinoderms without laterally directed arm-like extensions) contains the subclasses Echinoidea (the sea urchins and sand dollars) and Holothuroidea (the sea cucumbers). Of these two subclasses, only the Echinoidea are useful fossils.
These are typical star-shaped free-moving echinoderms in which the body is divided into a central disk and radiating arms.
This class contains the starfishes which, although not common fossils, illustrate well the typical echinoderm characteristics (Pl. 35). Fossil starfishes have been found sparingly in certain formations in Texas, but well-preserved specimens are quite rare. However, excellently preserved starfishes have been found in slabs of Cretaceous limestones from central and north-central Texas.
The ophiuroids are echinoderms with a well-defined central disk and five long, slender, whip-like arms. They have been called brittle stars because of their ability to shed their arms when they are disturbed. Their long, slender, snake-like arms have also resulted in their being called serpent stars. Ophiuroids range from Ordovician to Recent in age, but because of the delicate nature of their bodies they are seldom found as fossils. Ophiuroid remains have been found in certain Mesozoic and Cenozoic rocks of Texas, but they consist largely of small segments of the arms or body fragments.
The echinozoans are a group of unattached echinoderms whose bodies consist of numerous calcareous plates and spines, but they do not possess the radiating arm-like extensions which characterize the asterozoans.
Echinoids are free-moving echinoderms with disk-shaped, heart-shaped, biscuit-shaped, or globular exoskeletons (Pl. 36). Modern representatives of this group include the familiar sea urchins, heart urchins, and the sand dollars.
Plate 35
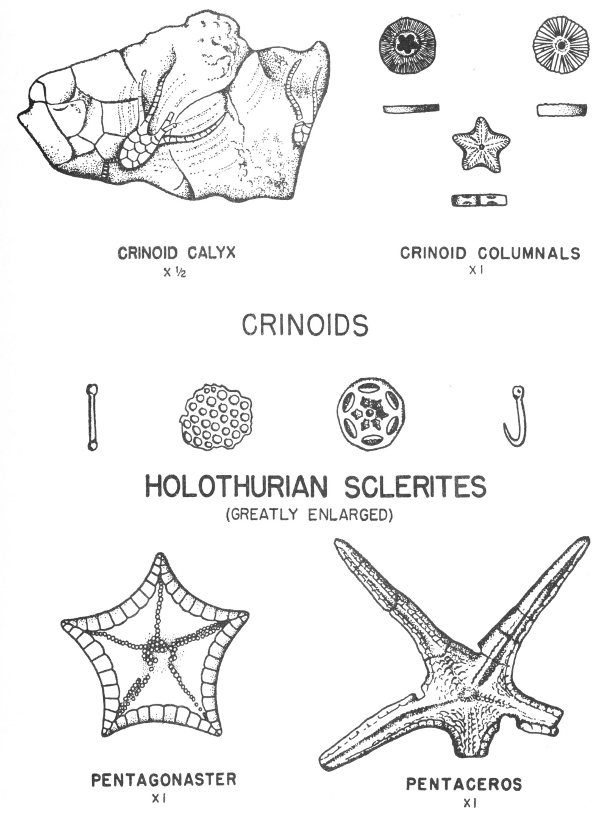
Plate 36
CRETACEOUS ECHINOIDS

The echinoid test (exoskeleton) is composed of many intricately fitting calcareous plates (Pl. 36) which enclose the animal’s soft parts. The exterior of the test is typically covered with large numbers of movable spines (Pl. 36) which vary greatly in size. These spines are of some aid in locomotion, support the skeleton of the animal, and provide a measure of protection from enemies.
The oldest known echinoids have been recorded from rocks of Ordovician age, but it was not until the Mesozoic that the group began to flourish. They were especially abundant during the Cretaceous and have been abundant and varied from that time until the present.
Echinoids are particularly numerous in many of the Lower Cretaceous formations of Texas where they are commonly found in an excellent state of preservation. Heart urchins and biscuit urchins may be found in large numbers in many areas of the State, and especially in areas where there are good exposures of fossiliferous Lower Cretaceous rocks.
Members of this class, commonly called sea cucumbers, have a rather elongate, sac-like, cucumber-shaped body and bear little resemblance to other members of the phylum Echinodermata. The sea cucumbers do not have a well-defined skeleton; rather the body is supported by many small, disconnected, calcareous plates or rods called ossicles or sclerites (Pl. 35). These minute structures are embedded in the leathery skin which covers the body of the sea cucumber and may be preserved as fossils. Such remains are locally abundant in certain formations in Texas, but because of their small size, scattered occurrence, and problems in classification, this group is of little use to most paleontologists.
Holothuroid body impressions have been reported from the Middle Cambrian, and sclerites from rocks as old as Mississippian.
The chordates are the most advanced of all animals and are characterized by the presence of a well-developed nervous system and a body supported by a bony or cartilaginous notochord and/or spinal column. In the higher chordates (the vertebrates) the notochord is normally replaced by bone, but in the lower chordates (for example, the graptolites) it remains in a cartilaginous condition.
The phylum Chordata contains only two subphyla of paleontological significance. These are the subphylum Hemichordata, composed of primitive chordates (including the graptolites which are important fossils), and the Vertebrata, which includes all animals with backbones.
The hemichordates are characterized by a well-defined notochord which runs the length of the body, but they do not possess a true backbone. Only one class, the Graptolithina, is of paleontological importance.
The graptolites are a group of extinct colonial animals which were very abundant during early Paleozoic time. They are characterized by a chitinous exoskeleton consisting of rows of cups or tubes which housed the living animal. These cups are attached to single or branching stalks (fig. 24) which in some forms were attached to sea weeds, rocks, or other foreign objects where they led a fixed existence. The stalks of the unattached graptolites grew on floats (fig. 24a) and these floating forms attained wide geographic distribution. It is also possible that some of the attached forms were fixed to floating objects, such as sea weed, and thus were distributed in this manner.
Previous classifications have recognized the graptolites as members of the phylum Coelenterata. As coelenterates they were assigned, at various times, to the classes Hydrozoa, Scyphozoa, and Graptozoa. In addition, they were also classified as bryozoans by certain of the early paleontologists. This publication, in keeping with recent changes in taxonomy, considers graptolites to be an extinct group of hemichordates. This classification is based upon research in which uncompressed 86 graptolites were etched out of chert and studied in great detail. Information derived from these relatively undistorted specimens indicates a much higher degree of body organization than was previously suspected, and as a result of these studies most paleontologists now consider graptolites to be some form of primitive chordate.
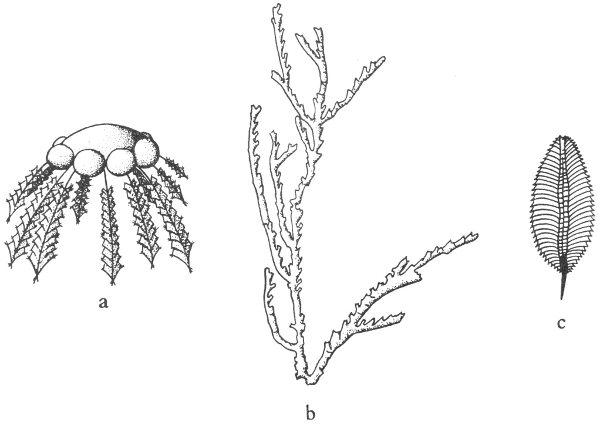
Fig. 24. Graptolites. (a) Diplograptus (×2). (b) Dendrograptus (×3). (c) Phyllograptus (×2).
The chitinous graptolite exoskeleton is commonly preserved as a flattened carbon residue; their remains may be locally abundant along the bedding planes of certain black or dark gray shales.
Graptolites are known from rocks that range from Cambrian to Mississippian in age, and they are among the most important guide fossils for Ordovician and Silurian rocks.
Graptolites have been reported from Cambrian rocks in central Texas and from the Ordovician of west Texas (fig. 24). The most abundant of these occur in certain Ordovician rocks in the Trans-Pecos area where they are common fossils in certain formations.
The vertebrates are the most advanced of all chordates. They are characterized by a skull and a bony or cartilaginous internal skeleton, with a vertebral column of bone or cartilage. This subphylum is commonly divided into two superclasses, the Pisces (the fishes and their relatives) and the Tetrapoda (the four-footed animals).
As mentioned earlier, most amateur collectors collect very few vertebrate remains, and for this reason this group is not discussed in detail. However, the more important vertebrate classes are briefly reviewed to enable the reader to have some understanding of this important group of animals. This part of the handbook will also serve as an introduction to some of the interesting and unusual, but now extinct, animals that have inhabited Texas in the geologic past. Among these animals are giant fishes, primitive amphibians, and many different types of dinosaurs. Included also are such unusual mammals as the giant ground sloths, saber-tooth cats, 87 mammoths, and mastodons, all of which are now extinct. The remains of these, and many other interesting extinct vertebrates, may be seen in the geological collections of the Texas Memorial Museum at Austin. Many of these displays are accompanied by drawings which depict the scientific restoration of the animal’s soft parts and show how the animal may have appeared in life.
The members of this superclass are commonly called fishes and are the simplest and most numerous of all vertebrates. They are aquatic, free-moving, cold-blooded (their blood maintains the temperature of the surrounding water), and breathe primarily by means of gills. However, some forms (the lungfishes) breathe by means of a lung developed from the air-bladder.
The most recent fish classification recognizes four major classes, the Agnatha (primitive jawless fishes), the Placodermi (armored fishes with primitive jaws), the Chondrichthyes (sharks and related forms with cartilaginous internal skeletons), and the Osteichthyes (true bony fishes).
Fishes belonging to this class are primitive, jawless, and represented by the living lampreys and hagfishes. The first agnathans appeared in the Ordovician and were armored by a bony covering on the front part of their bodies. These primitive fishes, called ostracoderms, are the earliest recorded fishes and, in addition, appear to be the first known vertebrate animals. The ostracoderms first appeared in late Ordovician time, increased in numbers in the Silurian, and were extinct by the end of the Devonian.
These are primitive jaw-bearing fishes, the majority of which were heavily armored (Pl. 37). The placoderms were shark-like in appearance, and some of them grew to be as much as 30 feet in length. Members of this class appeared first in the Devonian and lasted into the Permian, at which time they became extinct. Placoderms are rare in Texas, but the fragmentary remains of these primitive fishes have been found in Devonian rocks in central Texas.
This class includes such modern forms as the sharks, rays, and skates. They are characterized by skeletons which are composed of cartilage and are very abundant in the marine waters of today. The earliest known representatives of this class are reported from rocks of Devonian age, and they have been relatively common up to the present time.
Shark teeth (Pl. 37) can be found in Texas in Pennsylvanian, Permian, Cretaceous, Paleocene, Eocene, and Miocene rocks. These are probably the most common vertebrate fossils to be found in Texas and are usually found in thin-bedded marine limestones or clays.
The Osteichthyes includes the true bony fishes, which are the most highly developed and abundant of all fishes. They possess an internal bony skeleton, well-developed jaws, an air-bladder, and, typically, an external covering of overlapping scales.
Included in this class are a primitive group of fishes called crossopterygians. These were very abundant in the Devonian and are believed to be the ancestors of the amphibians. The modern lungfishes also belong to the class Osteichthyes, and these primitive fishes, which are now found only in Australia, South America, and Africa, breathe by means of gills and lungs which have been developed from the air-bladder. Although not abundant as fossils, the remains of these specialized fishes have added much to present knowledge concerning the development of certain of the higher vertebrates.
The remains of bony fishes have been collected at many localities in Texas, and fossils of this type have been found primarily in rocks of Cretaceous age but have been reported from other rocks as well. Fish fossils are more commonly found in the form of teeth (Pl. 37), vertebrae, scales, and an occasional well-preserved skeleton.
Plate 37
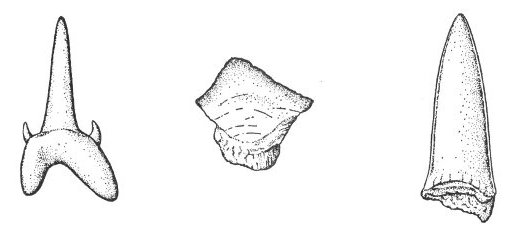
SHARK TEETH × 1
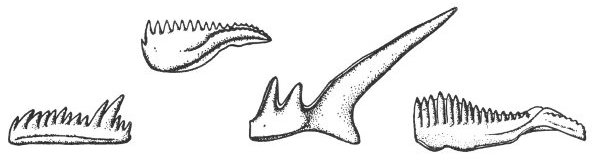
CONODONTS
(GREATLY ENLARGED)
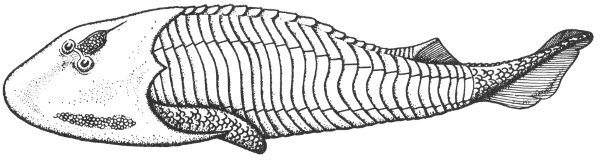
PRIMITIVE ARMORED FISH
PLACODERM (DEVONIAN)
× ½
Conodonts (Pl. 37) are small, amber-colored, tooth-like fossils which are believed to represent the teeth of some type of extinct fish. Although geologists do not know a great deal about the origin of these strange fossils they are of value in micropaleontology. Conodonts have been reported from several Paleozoic formations in Texas and are useful guide fossils in some areas.
The tetrapods are the most advanced chordates and are typified by the presence of lungs, a three- or four-chambered heart, and paired appendages. Included here are the classes Amphibia (frogs, toads, and salamanders), Reptilia (lizards, snakes, turtles, and the extinct dinosaurs), Aves (birds), and Mammalia (including the mammals, such as men, dogs, whales, etc.).
The amphibians were the earliest developed four-legged animals and are represented by such living forms as the toads, frogs, and salamanders. Amphibians are cold-blooded animals that primarily breathe by lungs and spend most of their life on land, but during their early stages of development they live in the water where they breathe by means of gills.
The amphibians apparently developed from the crossopterygian fishes during late Devonian time and were relatively abundant in the Pennsylvanian, Permian, and Triassic.
Amphibian remains in Texas are confined largely to lower Paleozoic and upper Mesozoic rocks. Numerous interesting and important discoveries of fossil amphibians have been made in north and west Texas where their remains (Pl. 40) have been collected in association with early types of reptiles. The areas where Permian red beds are exposed in Archer and Baylor counties and where Triassic red beds are exposed from Big Spring north along the edge of the High Plains have furnished most of these specimens.
The reptiles have become adapted to permanent life on land and need not rely on an aquatic environment. They are cold-blooded and are normally characterized by a scaly skin. Reptiles have been much more abundant in the past than they are today, and they assumed many different shapes and sizes in the geologic past. Modern classifications recognize a large number of reptilian groups, but only the more important of these are briefly reviewed here.
These were a group of primitive reptiles which, although retaining some amphibian characteristics, became adapted to an exclusively land-dwelling existence. The cotylosaurs lived during the Pennsylvanian and Permian and apparently became extinct sometime during the late Permian. Cotylosaurs (Pl. 40) are well known from the Permian of north Texas.
These are reptiles in which the body is more or less completely enclosed by bony plates. This group is first known as fossils from late Triassic rocks of Europe, and modern representatives of the group include the turtles and tortoises. Fragmentary remains of turtle shells are among the most common vertebrate fossils found in the Tertiary. Some of the late Tertiary land tortoises were 3 to 4 feet long. The earliest known turtles in Texas have been found in Cretaceous rocks.
The pelycosaurs were a group of late Paleozoic reptiles some of which were characterized by the presence of a tall fin on their back (Pl. 40). The fossils of these unusual creatures are well known from the Permian red beds of north-central Texas.
The therapsids were a mammal-like group of reptiles which were well developed for a terrestrial existence. Although the remains of these primitive reptiles are not particularly important fossils, study of the therapsids has provided much valuable information about the origin of the mammals. Members of this group appeared first in the middle Permian and persisted until the middle Jurassic, but therapsid remains have not been reported from Texas.
Plate 38
Comparison of the dinosaurs. Reproduced with permission of Dr. J. W. Dixon, Jr., and Geology Department, Baylor University, Waco, Texas.
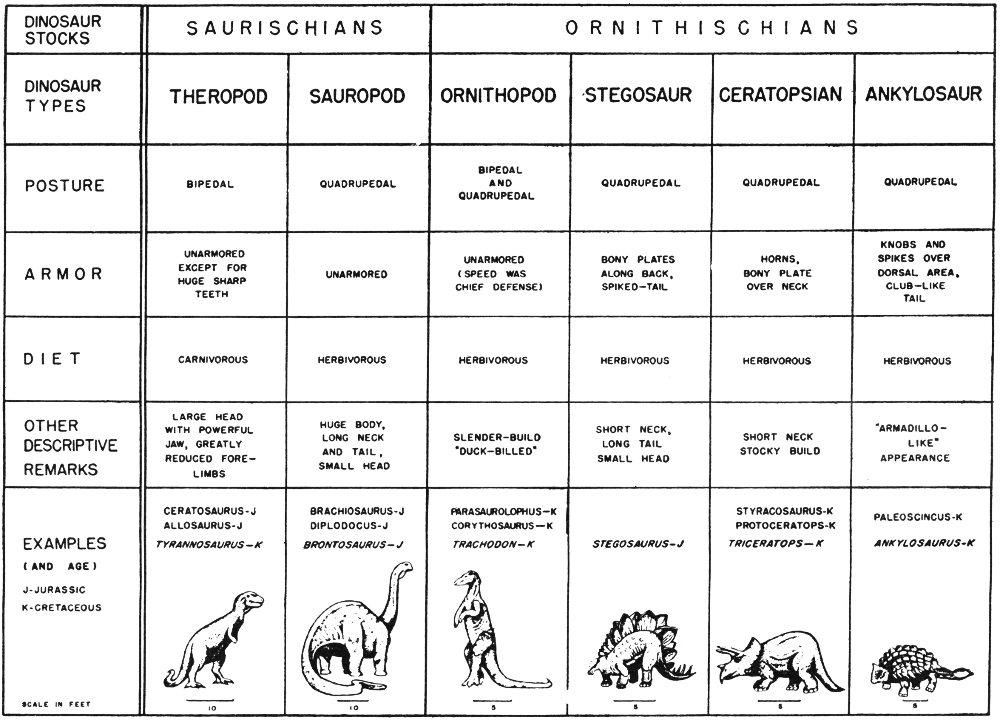
| DINOSAUR STOCKS | SAURISCHIANS | ORNITHISCHIANS | |||||
|---|---|---|---|---|---|---|---|
| DINOSAUR TYPES | THEROPOD | SAUROPOD | ORNITHOPOD | STEGOSAUR | CERATOPSIAN | ANKYLOSAUR | |
| POSTURE | BIPEDAL | QUADRUPEDAL | BIPEDAL AND QUADRUPEDAL | QUADRUPEDAL | QUADRUPEDAL | QUADRUPEDAL | |
| ARMOR | UNARMORED EXCEPT FOR HUGE SHARP TEETH | UNARMORED | UNARMORED (SPEED WAS CHIEF DEFENSE) | BONY PLATES ALONG BACK, SPIKED-TAIL | HORNS, BONY PLATE OVER NECK | KNOBS AND SPIKES OVER DORSAL AREA, CLUB-LIKE TAIL | |
| DIET | CARNIVOROUS | HERBIVOROUS | HERBIVOROUS | HERBIVOROUS | HERBIVOROUS | HERBIVOROUS | |
| OTHER DESCRIPTIVE REMARKS | LARGE HEAD WITH POWERFUL JAW, GREATLY REDUCED FORE-LIMBS | HUGE BODY, LONG NECK AND TAIL, SMALL HEAD | SLENDER-BUILD, “DUCK-BILLED” | SHORT NECK, LONG TAIL, SMALL HEAD | SHORT NECK, STOCKY BUILD | “ARMADILLO-LIKE” APPEARANCE | |
| EXAMPLES (AND AGE) J-JURASSIC K-CRETACEOUS | CERATOSAURUS-J ALLOSAURUS-J TYRANNOSAURUS-K | BRACHIOSAURUS-J DIPLODOCUS-J BRONTOSAURUS-J | PARASAUROLOPHUS-K CORYTHOSAURUS-K TRACHODON-K | STEGOSAURUS-J | STYRACOSAURUS-K PROTOCERATOPS-K TRICERATOPS-K | PALEOSCINCUS-K ANKYLOSAURUS-K | |
| SCALE IN FEET | 10 | 10 | 5 | 5 | 5 | 5 | |
Plate 39
Comparison of Mesozoic flying and swimming reptiles. Reproduced with permission of Dr. J. W. Dixon, Jr., and Geology Department, Baylor University, Waco, Texas.
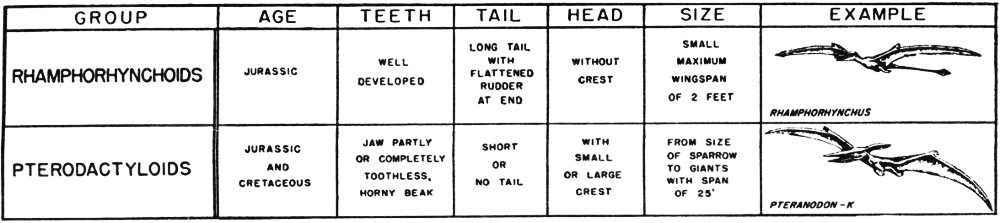
FLYING REPTILES—PTEROSAURS
| GROUP | AGE | TEETH | TAIL | HEAD | SIZE | EXAMPLE |
|---|---|---|---|---|---|---|
| RHAMPHORHYNCHOIDS | JURASSIC | WELL DEVELOPED | LONG TAIL WITH FLATTENED RUDDER AT END | WITHOUT CREST | SMALL MAXIMUM WINGSPAN OF 2 FEET | RHAMPHORHYNCHUS |
| PTERODACTYLOIDS | JURASSIC AND CRETACEOUS | JAW PARTLY OR COMPLETELY TOOTHLESS, HORNEY BEAK | SHORT OR NO TAIL | WITH SMALL OR LARGE CREST | FROM SIZE OF SPARROW TO GIANTS WITH SPAN OF 25′ | PTERANODON-K |
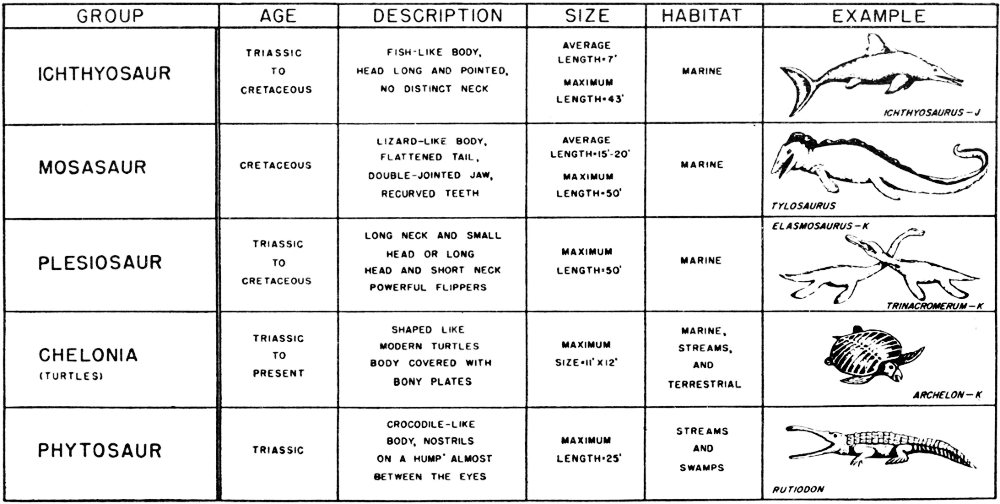
MESOZOIC SWIMMING REPTILES
| GROUP | AGE | DESCRIPTION | SIZE | HABITAT | EXAMPLE |
|---|---|---|---|---|---|
| ICHTHYOSAUR | TRIASSIC TO CRETACEOUS | FISH-LIKE BODY, HEAD LONG AND POINTED, NO DISTINCT NECK | AVERAGE LENGTH = 7′ MAXIMUM LENGTH = 43′ | MARINE | ICHTHYOSAURUS-J |
| MOSASAUR | CRETACEOUS | LIZARD-LIKE BODY, FLATTENED TAIL, DOUBLE-JOINTED JAW, RECURVED TEETH | AVERAGE LENGTH = 15′-20′ MAXIMUM LENGTH = 50′ | MARINE | PYLOSAURUS |
| PLESIOSAUR | TRIASSIC TO CRETACEOUS | LONG NECK AND SMALL HEAD OR LONG HEAD AND SHORT NECK, POWERFUL FLIPPERS | MAXIMUM LENGTH = 50′ | MARINE | ELASMOSAURUS-K TRINACROMERUM-K |
| CHELONIA (TURTLES) | TRIASSIC TO PRESENT | SHAPED LIKE MODERN TURTLES, BODY COVERED WITH BONY PLATES | MAXIMUM SIZE = 11′ × 12′ | MARINE, STREAMS, AND TERRESTRIAL | ARCHELON-K |
| PHYTOSAUR | TRIASSIC | CROCODILE-LIKE BODY, NOSTRILS ON A “HUMP” ALMOST BETWEEN THE EYES | MAXIMUM LENGTH = 25′ | STREAMS AND SWAMPS | RUTIODON |
Plate 40
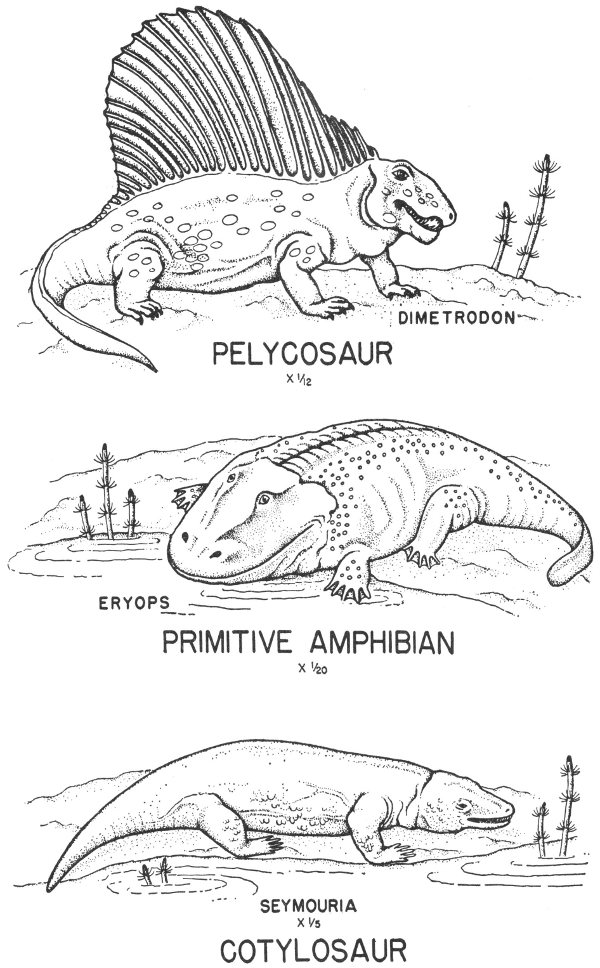
Plate 41
SWIMMING REPTILES
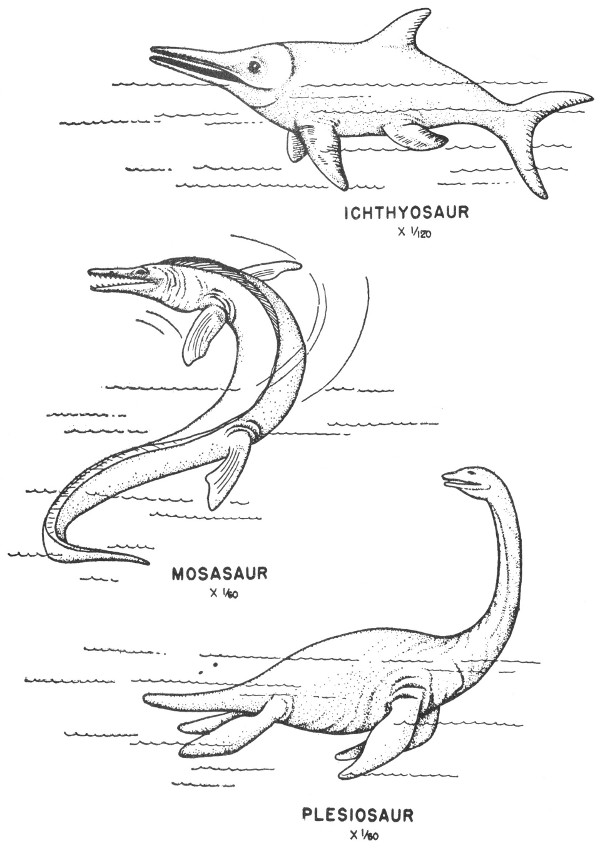
Plate 42
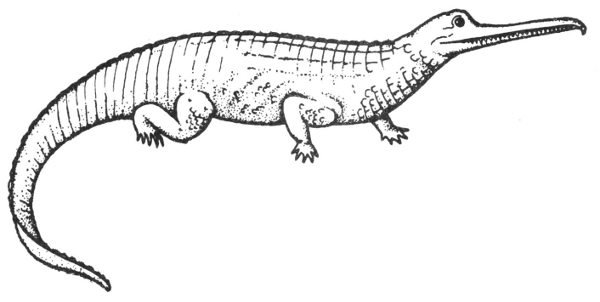
CROCODILE-LIKE REPTILE
× ¹/₅₀
PHYTOSAUR
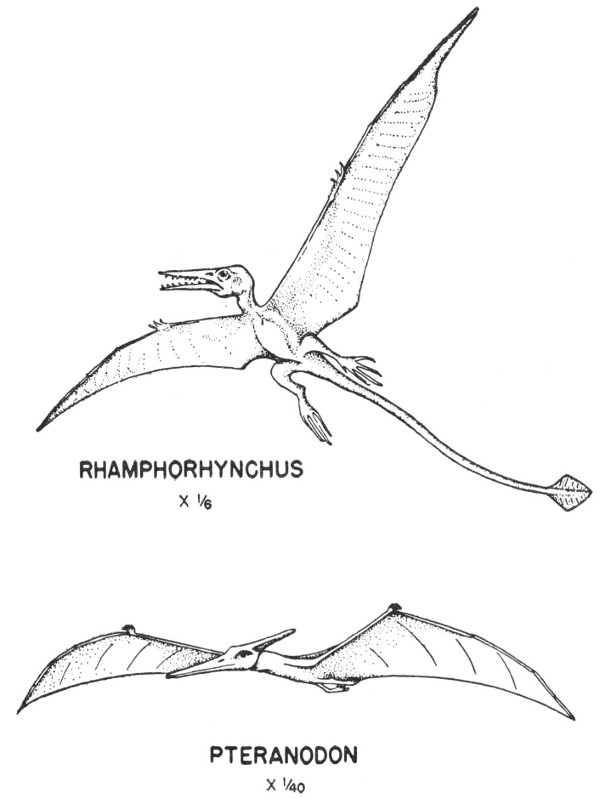
FLYING DINOSAURS
These were extinct, short-necked, marine reptiles that were fish-like in appearance. Ichthyosaurs resemble the modern dolphins, and some of them attained lengths of 25 to 40 feet (Pl. 41), though the average was much less. The group is known from rocks ranging from middle Triassic to late Cretaceous in age.
The mosasaurs are another group of extinct marine lizards which lived in Cretaceous seas. Some of these great reptiles grew to be as much as 50 feet long, and their great gaping jaws were filled with many sharp recurved teeth (Pl. 41). Mosasaurs were present in the great Cretaceous seas which covered many parts of Texas, and their remains have been reported from both north and central Texas. One such skeleton was found near Austin, and its skull is on display in the Texas Memorial Museum.
The plesiosaurs were marine reptiles which were characterized by a broad turtle-like body, paddle-like flippers, and a long neck and tail (Pl. 41). These reptiles were not as streamlined or well equipped for swimming as the ichthyosaurs or mosasaurs, but the long serpent-like neck was probably very useful in helping the reptile catch fish and other small animals for food. Plesiosaur remains range from middle Triassic to late Cretaceous in age, and they have been found in Cretaceous rocks in Texas. A short-necked plesiosaur which was collected from Upper Cretaceous rocks near Waco is on display in the Strecker Museum at Baylor University in Waco.
The phytosaurs were a group of crocodile-like reptiles which ranged from 6 to 25 feet in length (Pl. 42). They resembled the crocodiles both in appearance and in their mode of life, but this similarity is only superficial, and the phytosaurs and crocodiles are two distinct groups of reptiles.
The phytosaurs are exclusively Triassic in age and their remains have been collected from Triassic rocks along the eastern margin of the High Plains of Texas.
These reptiles adapted themselves to the same type habitat that was occupied by the phytosaurs, which preceded them. Crocodiles and alligators were much larger and more abundant during Cretaceous and Cenozoic time than they are today; the crocodiles first appeared in the Cretaceous and the alligators in the Tertiary. The remains of both crocodiles and alligators have been found in Texas, and one such crocodile (Phobosuchus) represents the remains of the largest crocodile yet discovered (Pl. 43). This specimen probably had an overall length of 40 to 50 feet, and its massive skull was 6 feet long and possessed exceptionally strong jaws. The remains of this great beast were collected from exposures of Upper Cretaceous rocks along the Rio Grande in Trans-Pecos Texas.
These were Mesozoic reptiles with bat-like wings supported by arms and long thin “fingers” (Pl. 42). The pterosaurs were well adapted to life in the air, and their light-weight bodies and wide skin-covered wings enabled them to soar or glide for great distances. The earliest known pterosaurs were found in lower Triassic rocks, and the group became extinct by the end of the Cretaceous. During this time certain of these creatures attained a wingspread of as much as 27 feet, but their bodies were small and light.
The collective term “dinosaurs” (meaning terrible lizards) has been given to that distinctive group of reptiles prominent in Mesozoic life for some 140 million years. In size, the dinosaurs ranged from as little as 1 foot to as much as 85 feet in length and from a few pounds to perhaps 45 tons in weight. Some were carnivorous (meat-eaters) but the majority were herbivorous (plant-eaters). Some were bipedal (walked on their hind-legs) while others were quadrupedal (walked on all fours), and although most of the dinosaurs were terrestrial in habitat, aquatic and semi-aquatic forms were also present.
According to the structure of their hip bones, the dinosaurs have been divided into two great orders. These are the Saurischia (forms with lizard-like pelvic girdle) and the Ornithischia (dinosaurs with a bird-like pelvic girdle).
Plate 43
Dr. Brown, R. T. Bird, and Dr. Schaikjer with the skull of Phobosuchus, an extinct crocodile from the Cretaceous of Trans-Pecos Texas.
Photograph courtesy of the American Museum of Natural History.
Dinosaurs belonging to this order were particularly abundant during the Jurassic and are characterized by hip bones that are similar to those of modern lizards. These dinosaurs were first discovered in rocks of Triassic age and did not become extinct until the end of the Cretaceous. The lizard-hipped reptiles are divided into two rather specialized groups of dinosaurs: the theropods (carnivorous bipedal dinosaurs that varied greatly in size) and the sauropods (herbivorous, quadrupedal, semi-aquatic, usually gigantic dinosaurs).
This type of saurischian dinosaur walked on bird-like hind limbs, and they were exclusively meat-eating forms, such as Allosaurus (Pl. 44) of Jurassic age. Some theropods were exceptionally large and were undoubtedly vicious beasts of prey. This assumption is borne out by such anatomical features as the small front limbs with long sharp claws for holding and tearing flesh, and the large strong jaws which were armed with numerous, sharp, dagger-like teeth. The largest of all known theropods was Tyrannosaurus rex which, when standing on his hind limbs, was almost 20 feet tall. Some individuals were as much as 50 feet long, and Tyrannosaurus is believed to have been among the most vicious animals to ever inhabit our earth. A cast of the skull of one of these great beasts is on display in the Texas Memorial Museum at Austin, and a Tyrannosaurus tooth has been found in the Big Bend National Park in Trans-Pecos Texas.
The sauropods were the largest of all dinosaurs, and some attained a length of 85 feet and probably weighed 40 to 50 tons (Brontosaurus, Pl. 44). They were primarily herbivorous dinosaurs which had become adapted to an aquatic or semi-aquatic type of existence and probably inhabited lakes, rivers, and swamps. The tracks of sauropod dinosaurs have been collected from Lower Cretaceous rocks in central Texas (Pl. 4) and Upper Cretaceous beds in Big Bend National Park in Trans-Pecos Texas.
The ornithischian, or bird-hipped dinosaurs, were herbivorous reptiles which were quite varied in form and size and appear to have been more highly developed than the saurischians. This order includes the duck-billed dinosaurs (ornithopods), the plate-bearing dinosaurs (stegosaurs), the armored dinosaurs (ankylosaurs), and the horned dinosaurs (ceratopsians). Ornithischian tracks are known from Cretaceous rocks in central and Trans-Pecos Texas.
These unusual dinosaurs were predominantly bipedal, semi-aquatic, and some (like the duck-billed dinosaurs) were highly specialized (Trachodon, Pl. 45).
The stegosaurs were herbivorous, quadrupedal ornithischians with large projecting plates down the back and heavy spikes on their tails. The Jurassic dinosaur Stegosaurus (Pl. 45) is most typical of the plate-bearing forms. This creature weighed about 10 tons, was some 30 feet long, and stood about 10 feet tall at the hips. Stegosaurus is characterized by a double row of large, heavy, pointed plates which run along the animal’s back. These plates begin at the back of the skull and stop near the end of the tail. The tail was also equipped with four or more long curved spikes which were probably used as a means of defense. The animal had a very small skull which housed a brain that was about the size of a walnut, and it is assumed that these, and all other dinosaurs, were of very limited intelligence.
Stegosaurus remains have not been discovered in Texas, but these, like certain other of the extinct vertebrates, are mentioned because of their interesting and unusual form.
The ankylosaurs were four-footed, herbivorous, Cretaceous dinosaurs which had relatively flat bodies. The skull and back of the animal were protected by bony armor, and the club-like tail was armed with spikes. Paleoscincus (Pl. 45), a typical ankylosaur, had large spines projecting from along the sides of the body and tail. The armored spiked back and the heavy club-like tail probably provided Paleoscincus with much-needed protection from the vicious meat-eating dinosaurs of Cretaceous time.
Plate 44
SAURISCHIAN DINOSAURS

Plate 45
ORNITHISCHIAN DINOSAURS
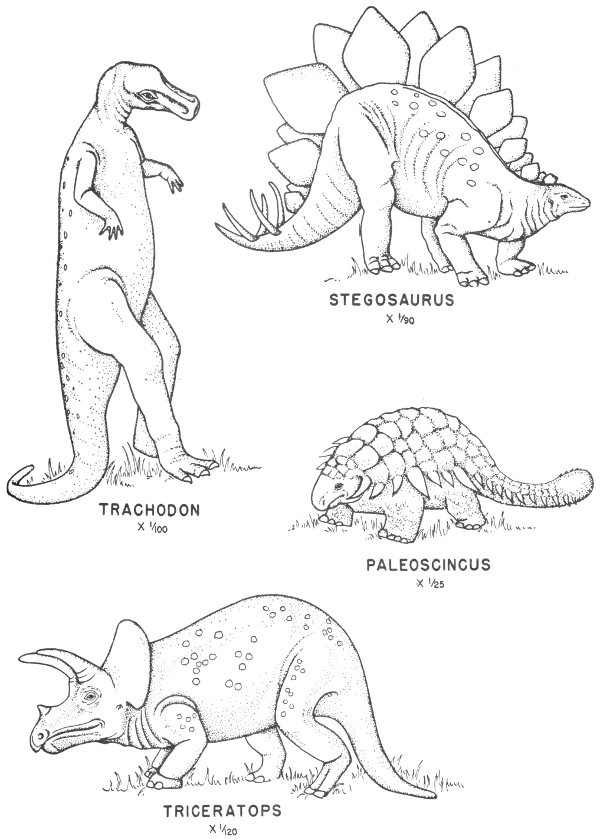
The ceratopsians, or horned dinosaurs, are another group of dinosaurs that are known only from rocks of Cretaceous age. These plant-eating dinosaurs possessed beak-like jaws, a bony neck frill which extended back from the skull, and one or more horns. Triceratops (Pl. 45) is the largest of the horned dinosaurs (some forms were as much as 30 feet long), and the skull measured 8 feet from the tip of the parrot-like beak to the back of the neck shield.
Because of the fragile nature of their bodies, birds are seldom found as fossils. In spite of this, however, some interesting and important fossil bird remains have been discovered.
The oldest known bird was found in Upper Jurassic rocks exposed in Germany. This primitive bird, named Archaeopteryx, is little more than a reptile with feathers. Archaeopteryx was a pigeon-sized creature which had scales as well as feathers, a lizard-like tail, a toothed beak, and other definitely reptilian characteristics.
During late Cretaceous time the birds underwent changes that resulted in forms similar to those that are living today, and most of the present-day birds had developed by the end of the Tertiary.
Although not commonly found, fossil birds have been recorded from certain of the Cenozoic rocks of Texas.
The mammals are animals that are born alive and fed with milk from the mother’s breast. They are warm-blooded, air-breathing, have a protective covering of hair, and are the most advanced of all vertebrates. The foregoing features are the more typical mammalian characteristics, but exceptions to these are found in certain mammals.
Mammals first appeared in the Jurassic and were probably derived from some form of mammal-like reptile. Although rare during the Mesozoic, mammals underwent rapid development and expansion during the Cenozoic, and during this era certain types of mammals became extremely large and assumed many bizarre shapes. The majority of these unusual forms lived but a short time but are well known from their fossils, and the remains of some of these animals which inhabited Texas during the Cenozoic may be seen in the Texas Memorial Museum at Austin.
Recent mammalian classification recognizes several subclasses and numerous orders and suborders, but the treatment of the mammals in a publication of this nature must of necessity be somewhat brief and no attempt at detailed classification is made.
The allotherians first appeared during the Jurassic and underwent considerable development in the late Cretaceous and early Tertiary. Included in this subclass are the multituberculates which are a group of small rodent-like animals that were probably the earliest of the herbivorous mammals. These animals were probably never very numerous, and they became extinct during the early part of Eocene time.
Members of this subclass are first known from rocks of Jurassic age, and they constitute the largest group of mammals that are living today. Therians undergo considerable development before they are born and at birth typically resemble the fully developed animal. This subclass has been divided into several orders but only the more important ones are discussed here.
The edentates are a rather primitive group of mammals which are represented by such living forms as the anteaters, tree sloths, and armadillos. Members of this group were common in the southern part of the United States in Pleistocene and Pliocene time, and fossil edentates have been reported from rocks of this age in Texas. One such form was Mylodon (Pl. 46), one of the extinct giant ground sloths. These huge sloths were quite heavy and some of them stood as much as 15 feet tall; these great creatures were the forerunners of the modern tree sloths of South America. The mounted skeleton of one of these giant ground sloths is displayed in the Texas Memorial Museum.
Plate 46
CENOZOIC MAMMALS
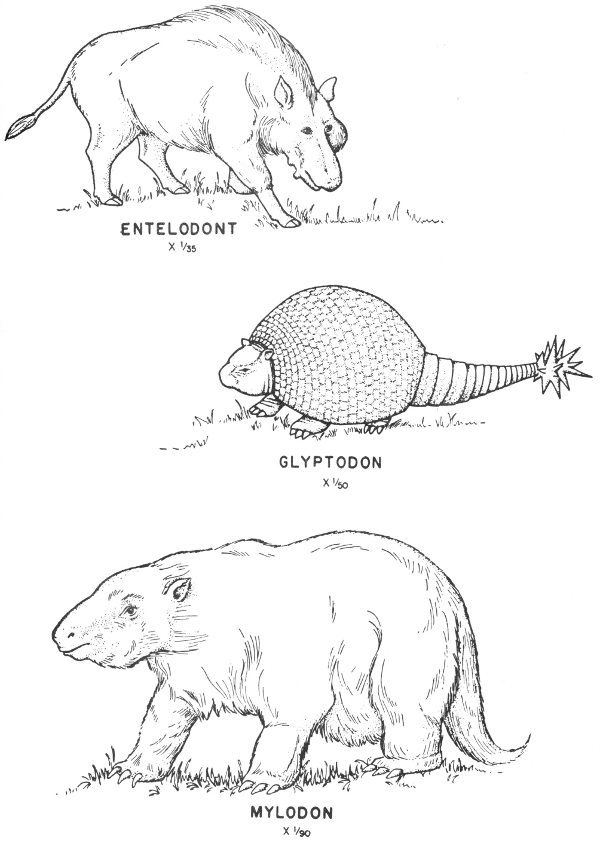
Another interesting representative of this order was the glyptodont. These peculiar mammals, which were ancestral to the present-day armadillos, developed at about the same time as the ground sloths. Glyptodon (Pl. 46), a typical glyptodont that has been reported from the Pleistocene of Texas, is quite characteristic of this group. This armadillo-like beast had a solid turtle-like shell that in some forms was as much as 4 feet high. From the front of the bone capped head to the tip of its tail, a large individual might be as much as 15 feet long. The thick heavy tail was protected by a series of bony rings, and in some species the end of the tail was developed into a bony heavily spiked club. The carapace (hard outer shell) of a large glyptodont is mounted at the Texas Memorial Museum.
Animals belonging to this order are called carnivores and are characterized by clawed feet and by teeth which are adapted for tearing and cutting flesh. The carnivores, or meat-eaters, were first represented by an ancient group of animals called creodonts, and this short-lived group first appeared in the Paleocene and were extinct by the end of the Eocene. They ranged from the size of a weazel to that of a large bear, and their claws were sharp and well developed. Their teeth, however, were not as specialized as those of modern carnivores, and the creodont brain was relatively small. It is assumed that these animals had a very low order of intelligence when compared to the more advanced carnivores of today.
These early meat-eaters were followed by more specialized carnivores which developed throughout Cenozoic time. Some examples of these are the saber-tooth cat Dinobastis (Pl. 47) and the dire wolf Canis diris (Pl. 47), both of which have been reported from the Texas Pleistocene. Some remains of these unusual forms, representing the cat and dog families, are on display at the Texas Memorial Museum.
Pantodonts, known also as amblypods, were primitive, hoofed, herbivorous animals. They were distinguished by a heavy skeleton, short stout limbs, and blunt spreading feet. The pantodonts appeared first during Paleocene time and had become extinct by the end of the Oligocene.
The members of this order are an extinct group of gigantic mammals commonly called uintatheres. Uintatherium (Pl. 48), which is typical of the group, had three pairs of blunt horns, and the males had dagger-like upper tusks. Some of the uintatheres were as large as a small elephant and stood as much as 7 feet tall at the shoulders. The size of the brain in relation to the size of the body suggests that these animals were not as intelligent as most mammals. Uintatheres are known from rocks ranging from Paleocene to Eocene in age. Uintathere remains have been reported from Big Bend National Park in Trans-Pecos Texas.
The earliest proboscideans, the elephants and their relatives, first appeared in the late Eocene of Africa and were about the size of a small modern elephant but had larger heads and shorter trunks. Proboscidean development is marked by an increase in size, change in skull and tooth structure, and elongation of the trunk. Two well-known fossil proboscideans are the mammoth and the mastodon, both of which inhabited Texas during Pleistocene time. The mastodons resembled the elephants, but the structure of their teeth was quite different (fig. 25). Moreover, the mastodon skull was lower than that of the elephant and the tusks were exceptionally large—some reaching a length of 9 feet.
Plate 47
CENOZOIC MAMMALS
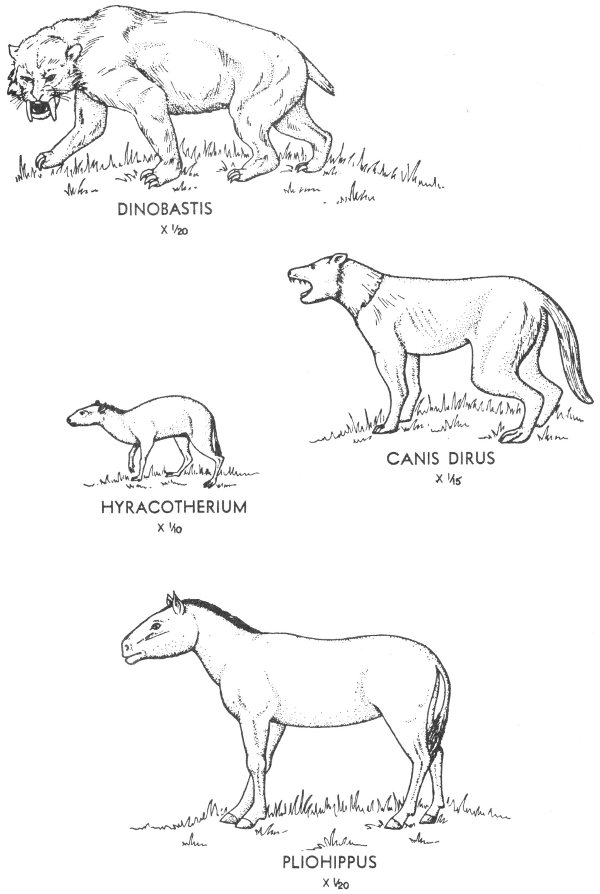
There were several types of mammoths, and the woolly mammoth is probably the best known. This animal lived until the end of the Pleistocene and, like the woolly rhinoceros discussed below, is known from ancient cave paintings and frozen remains. Information gathered from these sources indicates that this great beast had a long coat of black hair with a woolly undercoat (Pl. 49).

Fig. 25. Sketches of Pleistocene (a) mastodon tooth (×⅙) and (b) mammoth tooth (×⅙).
During the Pleistocene, mammoths were widespread over the United States, and their remains are abundant in many stream deposits of this age. Proboscidean bones have been reported from Pleistocene rocks in many parts of Texas, where they are commonly found in sand and gravel pits.
The perissodactyls, or odd-toed animals, are mammals in which the central toe on each limb is greatly enlarged. Modern representatives include the horses, rhinoceroses, and tapirs. Extinct members of the Perissodactyla include the titanotheres, chalicotheres, and baluchitheres, all of which grew to tremendous size and took on many unusual body forms.
One of the first perissodactyls was Hyracotherium (also called Eohippus), which is the earliest known horse (Pl. 47). This small animal, whose remains have been found in Big Bend National Park, was about 1 foot high and his teeth indicate a diet of soft food. Following the first horse, there is a long series of fossil horses which provide much valuable information on the history of this important group of animals.
The record of the development of the horse is well represented in Texas, and the bones and teeth of fossil horses are common in certain parts of the State. Fossils of this type have been reported from the Tertiary of the Trans-Pecos, Gulf Coastal Plain, and High Plains regions of Texas, and the teeth of Pleistocene horses have been found in sand and gravel pits in many parts of the State. Horse teeth (fig. 26) are particularly useful fossils as they may be accurately identified and used to determine the age of the rocks in which they are found.
This group of odd-toed mammals appeared first in the Eocene, at which time they were about the size of a sheep. By Middle Oligocene time they had increased to gigantic proportions but still had a small and primitive brain. Brontotherium (Pl. 48) was slightly rhinoceros-like in appearance and is believed to be the largest land animal that ever inhabited the North American continent. This animal was about 8 feet tall at the shoulders; a large bony growth protruded from the skull and this was extended into a flattened horn, which was divided at the top.
Plate 48
TERTIARY MAMMALS
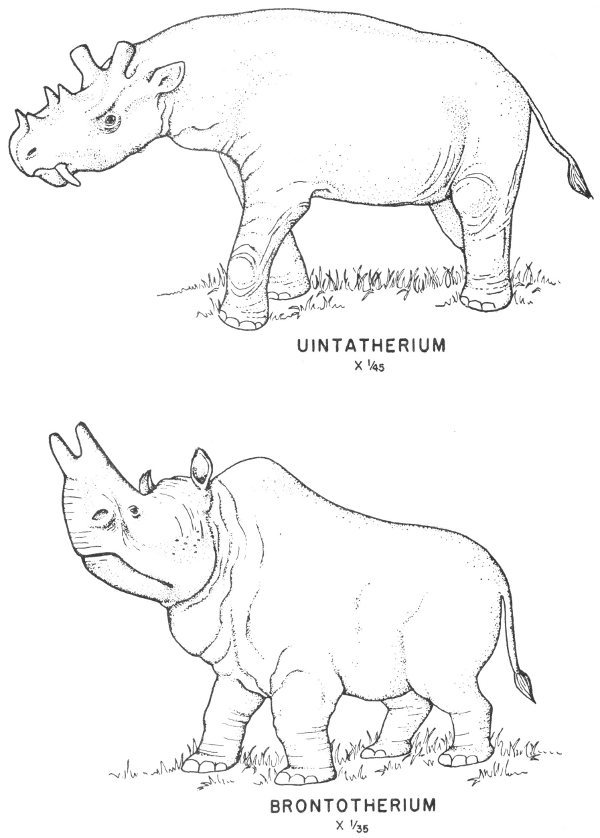
Although the titanotheres underwent rapid development during the early Tertiary, these huge beasts became extinct during the middle of the Oligocene epoch. Titanothere remains have been reported from the Trans-Pecos region of Texas.
The chalicotheres were in some ways like the titanotheres, but they also exhibited many peculiarities of their own. The head and neck of Moropus, a typical chalicothere, were much like that of a horse, but the front legs were longer than the hind legs, and the feet resembled those of a rhinoceros except that they bore long claws instead of hoofs. The chalicotheres lived in North America from Miocene until Pleistocene time but were probably never very numerous, and their remains have not yet been discovered in Texas.
The rhinoceroses are also odd-toed animals, and there are many interesting and well-known fossils in this group. The woolly rhinoceros (Pl. 49) was a Pleistocene two-horned form that ranged from southern France to northeastern Siberia. The woolly rhinoceros is well known from complete carcasses recovered from the frozen tundra of Siberia and from remains that were found preserved in an oil seep in Poland. These unusual specimens plus cave paintings made by early man have given a complete and accurate record of this creature. Although the woolly rhinoceros has not been reported from Texas, other fossil rhinoceroses have been found in the High Plains and Gulf Coastal Plain of Texas. These fossils have been found in rocks ranging from Middle Oligocene to late Pliocene in age.
Baluchitherium, the largest land mammal known to science, was a hornless rhinoceros that lived in late Oligocene and early Miocene time. This immense creature measured approximately 25 feet from head to tail, stood almost 18 feet high at the shoulder, and must have weighed many tons. Remains of these creatures have not been discovered in North America, and they appear to have been restricted to Central Asia.
The artiodactyls are the even-toed hoofed mammals and include such familiar forms as pigs, camels, deer, goats, sheep, and hippopotamuses. This is a large and varied group of animals, but the basic anatomical structure of the limbs and teeth show well the relationship between the different forms. Artiodactyls are abundant fossils in rocks ranging from Eocene to Pleistocene in age and are common in rocks of this age in Texas.
These giant pig-like artiodactyls lived during Oligocene and early Miocene time and were distinguished by a long heavy skull that held a relatively small brain. The face was marked by large knobs which were located beneath the eyes and on the underside of the lower jaw, and although these knob-like structures were blunt they had the appearance of short horns. Certain of these giant swine attained a height of 6 feet at the shoulders and had skulls that measured 3 feet in length (Pl. 46). Entelodont remains have been found in the Miocene of the Texas Coastal Plain.
The first known camels have been reported from rocks of upper Eocene age, and these small forms underwent considerable specialization of teeth and limbs as they developed in size. Many of the camels that lived during the middle Cenozoic had long legs which were well adapted to running and long necks which would have allowed the animals to browse on the leaves of tall trees.
The earliest known Texas camels were found in rocks of Oligocene age, and camels, like horses, must have been abundant in Texas during the Pleistocene for their fossilized remains are common in many parts of the State.
Plate 49
CENOZOIC MAMMALS

The following books are recommended for the reader who wants to know more about fossils and fossil collecting. The publications listed below cover various phases of historical geology and paleontology and range from children’s books to the more technical publications of the professional paleontologist. This list, however, is by no means all-inclusive and many other interesting and useful publications are available.
Dunbar, C. O. (1959) Historical geology, John Wiley and Sons, New York.
College-level text, well written and well illustrated.
Moore, R. C. (1958) Introduction to historical geology, McGraw-Hill Book Co., New York.
College-level presentation of earth history. Many illustrations of fossils.
Moore, Ruth (1953) Man, time, and fossils, Alfred Knopf, New York.
A readable account of fossils and their development throughout geologic time.
Panghorn, M. W., Jr. (1957) Earth for the layman, American Geological Institute, Washington, D. C.
Contains many valuable references.
Raymond, P. E. (1950) Prehistoric life, Harvard University Press, Cambridge, Mass.
College-level text.
Richards, H. G. (1953) Record of the rocks, Ronald Press, New York.
College-level earth history text.
Simpson, G. G. (1953) Life of the past, Yale University Press, New Haven, Conn.
Thorough, yet readable, introduction to paleontology.
Stirton, R. A. (1959) Time, life, and man: the fossil record, John Wiley and Sons, New York.
An introductory college text, most of which is of interest to adult level general readers.
Note: See also sections on Paleontology and Fossils in Encyclopedia Americana, Encyclopaedia Britannica, and others.
Andrews, R. C. (1953) All about dinosaurs, Random House, New York.
Interesting and readable dinosaur book for junior high and high-school age.
Andrews, R. C. (1956) All about strange beasts of the past, Random House, New York.
Interesting and easy to read, this book deals largely with extinct and unusual mammals (junior high and high school).
Colbert, E. H. (1945) The dinosaur book, American Museum of Natural History, New York.
A classic among “popular” dinosaur books. For all age levels.
Colbert, E. H. (1957) Dinosaurs, American Museum of Natural History, New York.
This little booklet provides a well-illustrated introduction to the dinosaurs. For high school and adult-level readers.
Dickinson, Alice (1954) First book of prehistoric animals, Franklin Watts, Inc., New York.
Easy to read, well-illustrated book for grade-school age.
Dunkle, D. H. (1957) The world of the dinosaurs, Smithsonian Institution, Washington, D. C.
An easy to understand, amply illustrated introduction to the dinosaurs (high school-adult level).
Fenton, C. L. (1937) Life long ago, The John Day Co., New York.
Very good for advanced grade and high-school age.
Heal, Edith (1930) How the world began, Thomas S. Rockwell Co., Chicago.
An account of the beginnings of life. For upper grade through high-school age.
Markman, H. C. (1954) Fossils, Denver Museum of Natural History, Denver, Colo.
A well-illustrated general survey of fossils. For adult-level readers.
Matthews III, W. H. (1962) Fossils: An introduction to prehistoric life, Barnes and Noble, Inc., New York, [“In preparation” at time of first printing of Guidebook No. 2.]
This publication contains many collecting aids and much background material for amateur collectors. Contains also a brief review of earth history.
Matthews III, W. H. (1963) Wonders of the dinosaur world, Dodd, Mead & Co., New York.
Well illustrated, non-technical presentation of dinosaurs. For junior-high and high-school teachers.
Parker, B. M. (1942) Stories read from the rocks, Basic Science Education Series, Row, Peterson and Co., Evanston, Ill.
Well written and colorfully illustrated. For advanced grades and junior high.
Parker, B. M. (1948) Animals of yesterday, Basic Science Education Series, Row, Peterson, and Co., Evanston, Ill.
Well written and colorfully illustrated. For advanced grades and junior high.
Shaver, R. H. (1959) Adventures with fossils, Geological Survey, Indiana Department of Conservation, Bloomington, Ind.
Collection hints and general information on fossils. Particularly for the lower grades.
Shuttlesworth, D. E. (1957) Real book of prehistoric life, Garden City Books, Garden City, N. Y.
Survey of prehistoric life. For grade and junior-high levels.
Brown, Vinson (1954) How to make a home nature museum, Little, Brown and Co., Boston.
Contains suggestions for collecting, mounting, and displaying fossils and other objects of nature.
Camp, C. L., and Hanna, G. D. (1937) Methods in paleontology, University of California Press, Berkeley.
Excellent discussion of collecting and preparation techniques.
Casanova, Richard (1957) An illustrated guide to fossil collecting, Natureograph Co., San Martin, Calif.
Has collecting hints and fossil localities for most of the States.
Collinson, C. C. (1959) Guide for beginning fossil hunters, Educational Series 4, Illinois State Geological Survey, Urbana.
Clearly written, well illustrated, particularly for the lower grades.
Goldring, Winifred (1950) Handbook of paleontology for beginners and amateurs, New York State Museum, Albany, N. Y.
A complete summary of paleontology. For the advanced collector.
La Rocque, A., and Marple, M. F. (1955) Ohio fossils, Ohio Division of Geological Survey, Bulletin 54, Columbus, Ohio.
Rather comprehensive treatment of the invertebrates with several useful keys for fossil identification.
Livingston, V. E., Jr. (1959) Fossils in Washington, Division of Mines and Geology, Department of Conservation, Olympia, Wash.
An introduction to the geology and fossils of Washington. Contains guide to collecting localities.
Simpson, B. W. (1958) Gem trails of Texas, Bessie W. Simpson, Granbury, Texas.
Field guide to Texas mineral, rock, and fossil locations. Contains numerous maps and well-described collecting localities.
Unklesbay, A. G. (1955) Common fossils of Missouri, University of Missouri Bulletin, Handbook 4, Columbia, Mo.
Written for the amateur; contains much general information of interest to the beginning collector.
Arnold, C. A. (1947) An introduction to paleobotany, McGraw-Hill Book Co., New York.
College-level textbook.
Beerbower, J. R. (1960) Search for the past, Prentice-Hall, Inc., Englewood Cliffs, N. J.
Good background text. Well illustrated. Has section on vertebrates.
Colbert, E. H. (1955) Evolution of the vertebrates, John Wiley and Sons, New York.
Comprehensive and technical treatment of vertebrate fossils.
Cushman, J. A. (1948) Foraminifera, their classification and economic use, Harvard University Press, Cambridge, Mass.
College-level text containing large numbers of descriptions and illustrations of foraminifera.
Easton, W. H. (1960) Invertebrate paleontology, Harper & Bros., Inc., New York.
College-level text. Good illustrations, useful for identification.
Fenton, C. L., and Fenton, M. A. (1958) The fossil book, Doubleday and Co., New York.
Comprehensive, easy-to-read, beautifully illustrated treatment of all types of fossils.
Jones, D. J. (1956) Introduction to microfossils, Harper and Brothers, New York.
College-level textbook with considerable information on collection, preparation, and the types of microfossils.
Moore, R. C., et al. (1953-1959) Treatise on invertebrate paleontology, Geological Society of America and University of Kansas, Lawrence, Kansas.
A technical reference for the more advanced collector. It is issued in several parts and contains latest classification.
Moore, R. C., Lalicker, C. G., and Fisher, A. G. (1953) Invertebrate fossils, McGraw-Hill Book Co., New York.
College-level reference with fine illustrations. Of value for purposes of identification.
Romer, A. S. (1945) Vertebrate paleontology, University of Chicago Press, Chicago.
A college-level textbook with numerous illustrations.
Shimer, H. W. (1933) Introduction to the study of fossils, The Macmillan Company, New York.
A relatively simple college-level presentation of plant and animal fossils.
Shimer, H. W., and Shrock, R. R. (1944) Index fossils of North America, John Wiley and Sons, New York.
Comprehensive survey of the more common fossils of North America. Useful to the advanced collector and a most useful aid for fossil identification.
Shrock, R. R., and Twenhofel, W. H. (1953) Principles of invertebrate paleontology, McGraw-Hill Book Co., New York.
Useful college-level reference for advanced collectors.
*Adkins, W. S. (1920) The Weno and Pawpaw formations of the Texas Comanchean: Univ. Texas Bull. 1856.
Descriptions and illustrations of many common Cretaceous fossils.
*Adkins, W. S. (1928) Handbook of Texas Cretaceous fossils: Univ. Texas Bull. 2838.
Lists all fossils described from the Texas Cretaceous prior to 1928, with many useful illustrations.
*Adkins, W. S., and Winton, W. M. (1919) Paleontological correlation of the Fredericksburg and Washita formations of north-central Texas: Univ. Texas Bull. 1945.
Contains descriptions and illustrations of many common Lower Cretaceous fossils of north-central Texas.
Clarke, W. B., and Twitchell, M. W. (1915) The Mesozoic and Cenozoic Echinodermata of the United States: U. S. Geological Survey Monograph 54, Washington, D. C.
A valuable guide to the Mesozoic and Cenozoic echinoderms of Texas.
*Frizzell, D. L. (1954) Handbook of Cretaceous Foraminifera of Texas: Univ. Texas, Bureau Econ. Geol. Rept. Inves. No. 22.
A technical, but invaluable aid in the study of Texas Cretaceous microfossils.
*Girard, R. M. (1959) Bibliography and index of Texas geology, 1933-1950: Univ. Texas Pub. 5910.
This valuable reference guide contains many references to Texas fossils. Note especially entries under Paleontology in the index.
Heuer, Edward (1958) Comments on the nomenclature revision of the Strawn and Canyon megafossil plates, in A guide to the Strawn and Canyon Series of the Pennsylvanian System in Palo Pinto County, Texas, An Occasional Publication of the North Texas Geological Society, Wichita Falls, Texas.
Contains illustrations and latest name changes of many of the more common Pennsylvanian fossils of north Texas.
*King, R. E. (1930) Geology of the Glass Mountains, Part II, Faunal summary and correlation of the Permian formations with description of Brachiopoda: Univ. Texas Bull. 3042.
Contains descriptions and illustrations of numerous brachiopods from the Glass Mountains of Trans-Pecos Texas.
*Lee, Wallace, et al. (1939) Stratigraphic and paleontologic studies of the Pennsylvanian and Permian rocks of north-central Texas: Univ. Texas Pub. 3801.
Contains an extensive faunal list and important collecting localities for Pennsylvanian invertebrates.
*Moore, R. C., and Jeffords, R. M. (1944) Description of lower Pennsylvanian corals from Texas and adjacent states: Univ. Texas Pub. 4401, pp. 77-208.
Describes and illustrates many of the more common Pennsylvanian corals.
*Plummer, F. B. (1943) The Carboniferous rocks of the Llano region of central Texas: Univ. Texas Pub. 4329.
Contains geologic map, locality data, and illustrations of many Carboniferous fossils.
*Plummer, F. B., and Moore, R. C. (1921) Stratigraphy of the Pennsylvanian formations of north-central Texas: Univ. Texas Bull. 2132.
Describes and illustrates many of the more common Pennsylvanian fossils of north-central Texas.
*Plummer, F. B., and Scott, Gayle (1937) Upper Paleozoic ammonites in Texas: Univ. Texas Bull. 3701, pt. 1.
*Renick, B. C., and Stenzel, H. B. (1931) The lower Claiborne of the Brazos River, Texas: Univ. Texas Bull. 3101, pp. 73-108.
Contains discussion and illustrations of many common Tertiary fossils.
Sellards, E. H. (1955) Texas through 250 million years: Museum Notes No. 4, Texas Memorial Museum, Austin.
This little booklet provides a short geologic history of Texas along with a review of oil in Texas.
*Sellards, E. H., Adkins, W. S., and Plummer, F. B. (1933) The geology of Texas, Vol. I, Stratigraphy: Univ. Texas Bull. 3232 (August 22, 1932).
This important publication will give the advanced collector much valuable information on the distribution of the rocks of Texas. Complete with geologic map.
Stanton, T. W. (1947) Studies of some Comanche pelecypods and gastropods: U. S. Geological Survey Prof. Paper 211, Washington, D. C.
Describes and illustrates most of the more common Lower Cretaceous pelecypods and gastropods of the State.
*Stenzel, H. B., Krause, E. K., and Twining, J. T. (1957) Pelecypoda from the type locality of the Stone City beds (Eocene) of Texas: Univ. Texas Pub. 5704.
Descriptions and illustrations of many of the more common Tertiary clams and oysters.
*Stephenson, L. W. (1941) The larger invertebrate fossils of the Navarro group of Texas: Univ. Texas Pub. 4101.
Contains descriptions of many common Upper Cretaceous invertebrates (exclusive of corals and crustaceans).
Stephenson, L. W. (1952) Larger invertebrate fossils of the Woodbine formation (Cenomanian) of Texas: U. S. Geological Survey Prof. Paper 242, Washington, D. C.
*Winton, W. M. (1925) The geology of Denton County: Univ. Texas Bull. 2544.
Illustrates and discusses the occurrence of many Cretaceous fossils.
*Winton, W. M., and Adkins, W. S. (1920) The geology of Tarrant County: Univ. Texas Bull. 1931.
Contains many illustrations of common north Texas Cretaceous fossils.
Amber—a hard, yellowish, translucent, fossilized plant resin.
Ammonite—ammonoid cephalopod with complexly wrinkled suture pattern; member of subclass Ammonoidea.
Anterior—front or fore.
Anus—the terminal opening of the alimentary canal, through which waste matter is discarded from the body.
Aperture—the opening of shells, cells, etc.
Aragonite—calcium carbonate (CaCO₃) crystallizing in a different form than calcite. In shells it is chalky and opaque; is less stable than calcite.
Archeozoic—the oldest known geological era; early Precambrian time.
Articulated—joined by interlocking processes or by teeth and sockets.
Asymmetrical—without or lacking symmetry.
Bilateral—pertaining to the two halves of a body as symmetrical and mirror images of each other.
Binomial nomenclature—system of scientific nomenclature requiring two names: generic and trivial.
Blastoid—stalked echinoderm with bud-like calyx usually consisting of 13 plates; member of class Blastoidea.
Brachiopod—bivalved marine invertebrate; member of phylum Brachiopoda.
Brackish—a mixture of salt and fresh waters.
Burrow—a hole in the ground, rock, wood, etc., made by certain animals for shelter or while gathering food.
Calcareous—composed of, or containing, calcium carbonate; limy.
Calcite—calcium carbonate (CaCO₃) crystallizing in a different form than aragonite. In shells it is translucent and more stable than aragonite.
Cambrian—the first (oldest) period of the Paleozoic era.
Calyx—in corals the bowl-shaped depression in the upper part of the skeleton; in stalked echinoderms that part of the body which contains most of the soft parts.
Caprinid—a Cretaceous pelecypod that is typically coiled in the form of a ram’s horn.
Carapace—the hard protective covering that forms the exoskeleton of many invertebrates; in arthropods it is usually chitinous or calcaro-chitinous.
Carbonization—the process of fossilization whereby organic remains are reduced to carbon or coal.
Cast—the impression taken from a mold.
Cenozoic—the latest era of geologic time, following the Mesozoic era and extending to the present.
Cephalon—the head; in trilobites the anterior body segment forming the head.
Cephalopod—marine invertebrate with well-defined head and eyes and with tentacles around the mouth; member of class Cephalopoda, phylum Mollusca; includes squids, octopuses, pearly nautilus.
Ceratite—an ammonoid cephalopod with suture composed of rounded saddles and jagged lobes; member of subclass Ammonoidea.
Chert—a cryptocrystalline variety of silica; flint is a variety of chert.
Chitin—a horn-like substance, found in the hard parts of many animals, such as beetles, crabs, etc.
Chitinous—composed of chitin.
Cirri—in crinoids, the jointed appendages which branch off the side of the stem or from the base of some crinoid stems.
Coelenterate—invertebrates characterized by a hollow body cavity, radial symmetry, and stinging cells; a member of phylum Coelenterata; includes jellyfishes, corals, sea anemones.
Colonial—in biology refers to the way in which some invertebrates live in close association with, and are more or less interdependent upon, each other; colonial corals, hydroids, etc.
Columella—a small column or central axis; in corals the small rod or axial pillar in the center of the corallite; in gastropods the solid or perforate pillar formed by the union of the successive coils of a conispiral shell.
Columnal—one of the disk-shaped segments of a crinoid stalk.
Concentric—having a common center, as circles; refers to shell markings that are parallel to shell margin.
Concretion—nodular or irregular masses in sedimentary rocks and usually formed around a central core, which is often a fossil.
Conical—cone-shaped.
Conodont—minute tooth-like fossils found in certain Paleozoic rocks; their origin is not definitely known, but they may have been part of some type of extinct fish.
Coral—bottom-dwelling marine invertebrate that secretes calcareous hard parts; member of class Anthozoa, phylum Coelenterata.
Corallite—the skeleton formed by an individual coral animal; may be solitary or form part of a colony.
Corallum—the skeleton of a coral colony.
Corona—crown; in echinoids the main part of the skeleton consisting of symmetrically arranged calcareous plates.
Coprolite—the fossil excrement of animals.
Correlation—the process of demonstrating that certain strata are closely related to each other or that they are stratigraphic equivalents.
Cretaceous—the third and last period of the Mesozoic era.
Cystoid—an extinct stemmed echinoderm with calyx composed of numerous irregularly arranged plates; member of class Cystoidea.
Dendritic—resembling a tree, branching.
Dentition—the system or arrangement of teeth peculiar to any given animal.
Devonian—the fourth oldest period of the Paleozoic era, follows the Silurian, precedes the Mississippian.
Dip—the angle of inclination which the bedding plane of rocks makes with a real or imaginary horizontal line.
Distillation—in fossils that process by which volatile organic matter is removed, leaving a carbon residue.
Dolomite—a mineral composed of calcium magnesium carbonate (CaMg(CO₃)₂).
Dorsal—pertaining to the back.
Echinoderm—a marine invertebrate with calcareous exoskeleton and usually exhibiting a five-fold radial symmetry; member of phylum Echinodermata; includes cystoids, blastoids, crinoids, starfishes, and sea urchins.
Echinoid—bottom-dwelling, unattached marine invertebrate with exoskeleton of calcareous plates covered by movable spines; member of class Echinoidea; sea urchins, heart urchins, biscuit urchins.
Endoskeleton—the internal supporting structure of an animal.
Eocene—the next to earliest of the Tertiary epochs, follows the Paleocene and precedes the Oligocene.
Equivalved—right and left valves subequal and (except for hinge structures) comprising mirror images of each other.
Evolution—a term applied to those methods or processes and to the sum of those processes whereby organisms change through successive generations.
Exoskeleton—an external skeleton, or hard covering for the protection of soft parts, particularly among invertebrates.
Fault—the displacement of rocks along a zone of fracture.
Fauna—an assemblage of animals (living or fossil) living in a given place at a given time.
Flank—the side or lateral portion of anything.
Flora—an assemblage of plants (living or fossil) living in a given place at a given time.
Fold—in brachiopods, a major rounded elevation of shell which affects both inner and outer shell surfaces.
Foramen—in brachiopods, the opening in the pedicle valve near the beak where the pedicle extends through the shell.
Foraminifer—a protozoan usually possessing a calcareous, perforated, chambered shell, but shell may be chitinous or agglutinated; a member of the order Foraminifera, phylum Protozoa.
Formation—a rock unit useful for mapping and distinguished primarily on the basis of lithologic characters.
Fossil—the remains or traces of organisms buried by natural causes and preserved in the earth’s crust.
Guide fossil—a fossil which, because of its limited vertical but wide horizontal distribution, is of value as a guide or index to the age of the rocks in which it is found.
Fossiliferous—containing fossilized organic remains.
Fusulinid—a spindle-shaped foraminifer: test shaped like a grain of wheat.
Gastrolith—highly polished well-rounded pebbles found associated with certain reptilian fossils; “stomach stones.”
Gastropod—a terrestrial or aquatic invertebrate, typically possessing a single-valved, calcareous, coiled shell; member of class Gastropoda, phylum Mollusca: snails and slugs.
Geologic age—the age of an object as stated in terms of geologic time (e.g., a Pennsylvanian fern, Cretaceous dinosaur).
Geologic map—map showing distribution of rock outcrops, structural features, mineral deposits, etc.
Geologic range—the known duration of an organism’s existence throughout geologic time (e.g., Cambrian to Recent for brachiopods).
Glauconite—a greenish mineral commonly formed in marine environments and essentially a hydrous silicate of iron and potassium.
Goniatite—an ammonoid cephalopod with suture composed of smooth saddles and simple angular lobes; member of subclass Ammonoidea.
Graptolite—an extinct, marine, colonial organism with chitinous hard parts; believed to belong to subphylum Hemichordata of phylum Chordata.
Guide fossil—see Fossil.
Habitat—the physical environment in which an organism lives.
Hinge-line—in brachiopods, the edge of the shell where the two valves articulate; in pelecypods, the dorsal margin of the valve which is in continual contact with the opposite valve.
Igneous rock—rocks which have solidified from lava or molten rock called magma.
Index fossil—see Fossil.
Inequivalved—opposite valves unlike in shape or size, or both.
Jurassic—second oldest period of the Mesozoic; follows the Triassic, precedes the Cretaceous.
Keel—a strong continuous ridge along the ventral side of ammonites.
Larva—the young form of some animals before they assume the mature shape.
Lateral—side or to the side.
Lithology—the study and description of rocks based on the megascopic (with the naked eye) examination of samples. Used also to refer to the texture and composition of any given rock sample.
Living chamber—in mollusks, that part of the shell which is occupied by the living animal.
Lobe—in cephalopods, the backward flexure of the suture or septum.
Longitudinal—in a direction parallel with the length.
Lophophore—in brachiopods, a tentacle-bearing appendage attached to the anterior surface of the mantle cavity.
Mantle—in mollusks and brachiopods, a layer of tissue containing cells that secrete the shell.
Meso-—a prefix signifying middle.
Mesozoic—that era of geologic time that precedes the Cenozoic and follows the Paleozoic.
Miocene—fourth oldest epoch of the Tertiary period; follows the Oligocene, precedes the Pliocene.
Mississippian—fifth oldest period of the Paleozoic: follows the Devonian, precedes the Pennsylvanian.
Multicellular—composed of more than one cell.
Nacreous—pearly.
Node—a knob.
-oid—a suffix meaning “in the form of.”
Oligocene—the third oldest epoch of the Tertiary period: precedes the Miocene, follows the Eocene.
Operculum—the lid or covering of the aperture of certain shells.
Oral—referring to the mouth or aperture.
Orbitoidids—foraminifers with large typically disk-shaped tests.
Ordovician—second oldest period of the Paleozoic era; follows the Cambrian, precedes the Silurian.
Ossicle—loosely used as a small plate.
Paleocene—oldest epoch of the Tertiary period; precedes the Eocene.
Paleozoic—that era of geologic time that follows Precambrian time and precedes the Mesozoic era.
Pedicle opening (pedicle foramen)—see Foramen.
Pelecypod—a bivalved aquatic invertebrate; member of class Pelecypoda, phylum Mollusca.
Pennsylvanian—the sixth oldest period of the Paleozoic era; follows the Mississippian, precedes the Permian.
Period—a division of geologic time (Pl. 1).
Periostracum—the horny outer covering or epidermis on shells.
Permian—seventh and last period of the Paleozoic.
Permineralization—that process by which mineral matter has been added to the original shell material by precipitation in the interstices rather than replacing the original shell material.
Phosphatic—containing or pertaining to phosphate minerals.
Phylum—one of the primary divisions of the animal or vegetable kingdoms.
Planispiral—shell coiled in one plane.
Pleistocene—earliest epoch of Quaternary period, Cenozoic era; follows Pliocene epoch of Tertiary period, precedes Recent epoch of Quaternary.
Pleural—referring to the side or ribs; in trilobites, refers to lateral portions of thorax and pygidium.
Pliocene—latest epoch of Tertiary period of Cenozoic era; follows Miocene epoch and precedes Pleistocene epoch of Quaternary period.
Polygonal—many sided or having many-sided plates.
Polyp—a many-tentacled aquatic coelenterate animal, typically cylindrical or cup-shaped, as in corals.
Porcelaneous—like porcelain.
Pore—a very small opening.
Posterior—situated behind; to the rear.
Precambrian—that portion of geologic time before the Cambrian; divided into Archeozoic era (Early Precambrian) and Proterozoic era (Late Precambrian).
Protero—combining form meaning fore, former, or anterior in time (Greek proteros, fore).
Proterozoic—youngest era of the Precambrian; follows the Archeozoic era and precedes the Cambrian period of the Paleozoic era.
Protista—the organic kingdom including the simplest of all one-celled organisms which possess various characters of both plants and animals; bacteria, algae, foraminifers, radiolarians.
Protoconch—in mollusks, the initial chamber of shell.
Pyrite—a hard, brass-yellow mineral composed of iron sulfide; “fool’s gold.”
Quaternary—the youngest period of the Cenozoic era, follows the Tertiary period.
Radial symmetry—see Symmetry.
Reef—a mound-like or ridge-like elevation of the sea bottom which almost reaches the surface of the water, composed primarily of organic material and commonly formed by reef-building animals, such as corals and oysters.
Replacement—type of fossilization whereby hard parts of organisms are removed by solution accompanied by almost simultaneous deposition of other substances in the resulting voids; mineralization.
Respiration—the process of oxygenation.
Rock—an aggregation of one or more minerals.
Rock-unit—divisions of rocks based on definite physical and lithologic characteristics and not defined on the basis of geologic time alone; groups, formations, members.
Rudistid—a Cretaceous pelecypod that does not exhibit the typical clam or oyster shape; many are cone-shaped, resembling corals.
Saddle—in cephalopods, the forward flexure (curved toward the aperture) of the suture or septum.
Scaphopod—an exclusively marine mollusk with a single-valved tusk-shaped shell; member of class Scaphopoda, phylum Mollusca.
Scavenger—an animal that feeds on organic refuse.
Sedentary—stationary in life, not moving from place to place.
Sediment—material that has been deposited by settling from a transportation agent such as water or air; typically composed of weathered rock fragments.
Sedimentary rock—rocks formed from the accumulation and lithification of sediments.
Segment—one of the parts into which a body naturally separates or is divided; for example, segments of arthropods or annelid worms.
Septal—pertaining to the septum.
Septum (plural, septa)—a dividing wall or partition; in fusulinids, a partition between chambers in the fusulinid shell; in corals, one of the radiating, longitudinal, calcareous plates located within the corallite; in cephalopods, the transverse partitions between the chambers.
Series—the rocks formed during an epoch; the time-stratigraphic term next in rank below a system.
Serrate—notched like a saw.
Sessile—animal attached to the sea floor more or less permanently.
Silica—an oxide of silicon (SiO₂).
Siliceous—containing or pertaining to silica.
Silicification—the process of combining or impregnating with silica.
Silurian—the third oldest period of the Paleozoic era; follows the Ordovician, precedes the Devonian.
Sinus—an elongate depression on brachiopod shells.
Siphuncle—in cephalopods, the segmented horny or calcareous tube which extends from the protoconch to the living chamber.
Slickensides—polished and grooved surfaces that are the result of two rock masses sliding past each other as in faulting.
Solitary—living alone; not part of a colony.
Species—one of the smaller natural divisions in classification.
Specific name—see Trivial name.
Spicule—a minute spike or dart, skeletal element in sponges and holothurians.
Stratum (plural, strata)—a single bed or layer of rock.
Strike—the direction of a real or imaginary line that is formed by the intersection of a bed or stratum with a horizontal plane; strike is perpendicular to the dip.
Subconical—less than conical in shape; almost a cone.
Suture—the line of junction between two parts; in crinoids, the line of junction between two plates; in gastropods, the line of junction of the whorls as seen on the exterior of the shell; in cephalopods, the line of junction between a septum and the shell wall.
Symmetry—orderly arrangement of parts of an object with reference to lines, planes, or points.
Bilateral symmetry—the symmetrical duplication of parts on each side of a vertical anterior-posterior plane.
Radial symmetry—the symmetrical repetition of parts around a common vertical dorso-ventrally disposed axis.
Pentamerous symmetry—symmetry arranged in a pattern of fives.
System—the rocks formed during a period; the time-stratigraphic term next in rank above a series.
Taxonomy—that branch of science that deals with classification, especially in relation to plants, animals, or fossils.
Tertiary—the oldest period of the Cenozoic era; follows the Cretaceous period of the Mesozoic and precedes the Quaternary period of the Cenozoic.
Test—the protective covering of some invertebrate animals.
Theca—a sheath or case; in coelenterates, the bounding wall at or near the margin of the exoskeleton; in echinoderms, the main body skeleton (or calyx) which houses the animal’s soft parts; in graptolites, any cup or tube of the colony.
Thorax—in trilobites, that part of the body between the cephalon and pygidium.
Time-unit—a portion of continuous geologic time (e.g., eras, periods, epochs, and ages).
Time-rock unit—same as time-stratigraphic unit.
Time-stratigraphic unit—term given to rock units with boundaries established by geologic time; strata deposited during definite portions of geologic time (e.g., systems, series, stages, etc.).
Topography—the physical features or configuration of a land surface.
Topographic map—a map showing the physical features of an area, especially the relief and contour of the land.
Transverse—at right angles to length.
Triassic—the youngest period of the Mesozoic era; follows the Permian period of the Paleozoic and precedes the Jurassic period of the Mesozoic.
Trilobite—an extinct marine arthropod having a flattened segmented body covered by a hardened dorsal exoskeleton divided into three lobes.
Trivial name—the Latinized name added to a generic name to distinguish the species; same as specific name.
Type locality—the geographic location at which a formation was first described and from which it was named; or from which the type specimen of a fossil species comes.
Type specimen—the individual or specimen on which the original designation of a species was established.
Umbilicus—an external depression or opening at the center of many loosely coiled shells; in gastropods it is usually located at the base of the shell; in cephalopods it is usually located laterally.
Umbo—the arched part of the valve near the beak in bivalve shells.
Unicellular—composed of one cell.
Valve—the one or more pieces comprising the shell of animals.
Variety—a subdivision of a species, designated by a third name when a variety is designated.
Ventral—pertaining to the abdomen; as opposed to dorsal, pertaining to the back.
Vertebrate—an animal having a backbone or spinal column.
Whorl—a single turn or volution of a coiled shell.
-zoic—combining form meaning “life” (Greek zoikos, life).
Zooecium (plural, zooecia)—tube or chamber occupied by an individual of the bryozoan colony; also called an autopore.
Page numbers in italics indicate illustrations.
A B C D E F G H I J K L M N O P Q R S T U V W X Y Z
End of the Project Gutenberg EBook of Texas Fossils: An Amateur Collector's
Handbook, by William H. Matthews III
*** END OF THIS PROJECT GUTENBERG EBOOK TEXAS FOSSILS ***
***** This file should be named 56315-h.htm or 56315-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/5/6/3/1/56315/
Produced by Stephen Hutcheson, MFR and the Online
Distributed Proofreading Team at http://www.pgdp.net
Updated editions will replace the previous one--the old editions will
be renamed.
Creating the works from print editions not protected by U.S. copyright
law means that no one owns a United States copyright in these works,
so the Foundation (and you!) can copy and distribute it in the United
States without permission and without paying copyright
royalties. Special rules, set forth in the General Terms of Use part
of this license, apply to copying and distributing Project
Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm
concept and trademark. Project Gutenberg is a registered trademark,
and may not be used if you charge for the eBooks, unless you receive
specific permission. If you do not charge anything for copies of this
eBook, complying with the rules is very easy. You may use this eBook
for nearly any purpose such as creation of derivative works, reports,
performances and research. They may be modified and printed and given
away--you may do practically ANYTHING in the United States with eBooks
not protected by U.S. copyright law. Redistribution is subject to the
trademark license, especially commercial redistribution.
START: FULL LICENSE
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full
Project Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project
Gutenberg-tm electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or
destroy all copies of Project Gutenberg-tm electronic works in your
possession. If you paid a fee for obtaining a copy of or access to a
Project Gutenberg-tm electronic work and you do not agree to be bound
by the terms of this agreement, you may obtain a refund from the
person or entity to whom you paid the fee as set forth in paragraph
1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this
agreement and help preserve free future access to Project Gutenberg-tm
electronic works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the
Foundation" or PGLAF), owns a compilation copyright in the collection
of Project Gutenberg-tm electronic works. Nearly all the individual
works in the collection are in the public domain in the United
States. If an individual work is unprotected by copyright law in the
United States and you are located in the United States, we do not
claim a right to prevent you from copying, distributing, performing,
displaying or creating derivative works based on the work as long as
all references to Project Gutenberg are removed. Of course, we hope
that you will support the Project Gutenberg-tm mission of promoting
free access to electronic works by freely sharing Project Gutenberg-tm
works in compliance with the terms of this agreement for keeping the
Project Gutenberg-tm name associated with the work. You can easily
comply with the terms of this agreement by keeping this work in the
same format with its attached full Project Gutenberg-tm License when
you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are
in a constant state of change. If you are outside the United States,
check the laws of your country in addition to the terms of this
agreement before downloading, copying, displaying, performing,
distributing or creating derivative works based on this work or any
other Project Gutenberg-tm work. The Foundation makes no
representations concerning the copyright status of any work in any
country outside the United States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other
immediate access to, the full Project Gutenberg-tm License must appear
prominently whenever any copy of a Project Gutenberg-tm work (any work
on which the phrase "Project Gutenberg" appears, or with which the
phrase "Project Gutenberg" is associated) is accessed, displayed,
performed, viewed, copied or distributed:
This eBook is for the use of anyone anywhere in the United States and
most other parts of the world at no cost and with almost no
restrictions whatsoever. You may copy it, give it away or re-use it
under the terms of the Project Gutenberg License included with this
eBook or online at www.gutenberg.org. If you are not located in the
United States, you'll have to check the laws of the country where you
are located before using this ebook.
1.E.2. If an individual Project Gutenberg-tm electronic work is
derived from texts not protected by U.S. copyright law (does not
contain a notice indicating that it is posted with permission of the
copyright holder), the work can be copied and distributed to anyone in
the United States without paying any fees or charges. If you are
redistributing or providing access to a work with the phrase "Project
Gutenberg" associated with or appearing on the work, you must comply
either with the requirements of paragraphs 1.E.1 through 1.E.7 or
obtain permission for the use of the work and the Project Gutenberg-tm
trademark as set forth in paragraphs 1.E.8 or 1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any
additional terms imposed by the copyright holder. Additional terms
will be linked to the Project Gutenberg-tm License for all works
posted with the permission of the copyright holder found at the
beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including
any word processing or hypertext form. However, if you provide access
to or distribute copies of a Project Gutenberg-tm work in a format
other than "Plain Vanilla ASCII" or other format used in the official
version posted on the official Project Gutenberg-tm web site
(www.gutenberg.org), you must, at no additional cost, fee or expense
to the user, provide a copy, a means of exporting a copy, or a means
of obtaining a copy upon request, of the work in its original "Plain
Vanilla ASCII" or other form. Any alternate format must include the
full Project Gutenberg-tm License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works
provided that
* You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is owed
to the owner of the Project Gutenberg-tm trademark, but he has
agreed to donate royalties under this paragraph to the Project
Gutenberg Literary Archive Foundation. Royalty payments must be paid
within 60 days following each date on which you prepare (or are
legally required to prepare) your periodic tax returns. Royalty
payments should be clearly marked as such and sent to the Project
Gutenberg Literary Archive Foundation at the address specified in
Section 4, "Information about donations to the Project Gutenberg
Literary Archive Foundation."
* You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or destroy all
copies of the works possessed in a physical medium and discontinue
all use of and all access to other copies of Project Gutenberg-tm
works.
* You provide, in accordance with paragraph 1.F.3, a full refund of
any money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days of
receipt of the work.
* You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project
Gutenberg-tm electronic work or group of works on different terms than
are set forth in this agreement, you must obtain permission in writing
from both the Project Gutenberg Literary Archive Foundation and The
Project Gutenberg Trademark LLC, the owner of the Project Gutenberg-tm
trademark. Contact the Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
works not protected by U.S. copyright law in creating the Project
Gutenberg-tm collection. Despite these efforts, Project Gutenberg-tm
electronic works, and the medium on which they may be stored, may
contain "Defects," such as, but not limited to, incomplete, inaccurate
or corrupt data, transcription errors, a copyright or other
intellectual property infringement, a defective or damaged disk or
other medium, a computer virus, or computer codes that damage or
cannot be read by your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium
with your written explanation. The person or entity that provided you
with the defective work may elect to provide a replacement copy in
lieu of a refund. If you received the work electronically, the person
or entity providing it to you may choose to give you a second
opportunity to receive the work electronically in lieu of a refund. If
the second copy is also defective, you may demand a refund in writing
without further opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO
OTHER WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT
LIMITED TO WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of
damages. If any disclaimer or limitation set forth in this agreement
violates the law of the state applicable to this agreement, the
agreement shall be interpreted to make the maximum disclaimer or
limitation permitted by the applicable state law. The invalidity or
unenforceability of any provision of this agreement shall not void the
remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in
accordance with this agreement, and any volunteers associated with the
production, promotion and distribution of Project Gutenberg-tm
electronic works, harmless from all liability, costs and expenses,
including legal fees, that arise directly or indirectly from any of
the following which you do or cause to occur: (a) distribution of this
or any Project Gutenberg-tm work, (b) alteration, modification, or
additions or deletions to any Project Gutenberg-tm work, and (c) any
Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of
computers including obsolete, old, middle-aged and new computers. It
exists because of the efforts of hundreds of volunteers and donations
from people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future
generations. To learn more about the Project Gutenberg Literary
Archive Foundation and how your efforts and donations can help, see
Sections 3 and 4 and the Foundation information page at
www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg Literary
Archive Foundation are tax deductible to the full extent permitted by
U.S. federal laws and your state's laws.
The Foundation's principal office is in Fairbanks, Alaska, with the
mailing address: PO Box 750175, Fairbanks, AK 99775, but its
volunteers and employees are scattered throughout numerous
locations. Its business office is located at 809 North 1500 West, Salt
Lake City, UT 84116, (801) 596-1887. Email contact links and up to
date contact information can be found at the Foundation's web site and
official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To SEND
DONATIONS or determine the status of compliance for any particular
state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations. To
donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic works.
Professor Michael S. Hart was the originator of the Project
Gutenberg-tm concept of a library of electronic works that could be
freely shared with anyone. For forty years, he produced and
distributed Project Gutenberg-tm eBooks with only a loose network of
volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as not protected by copyright in
the U.S. unless a copyright notice is included. Thus, we do not
necessarily keep eBooks in compliance with any particular paper
edition.
Most people start at our Web site which has the main PG search
facility: www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.
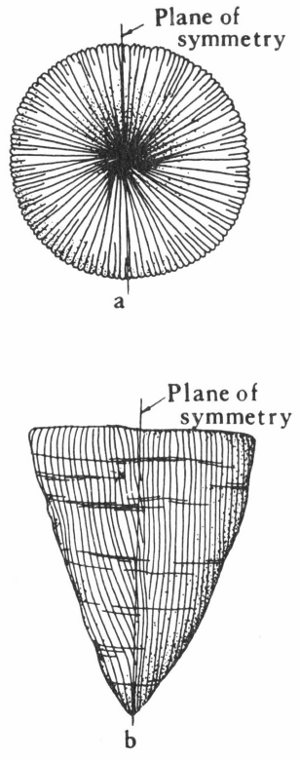

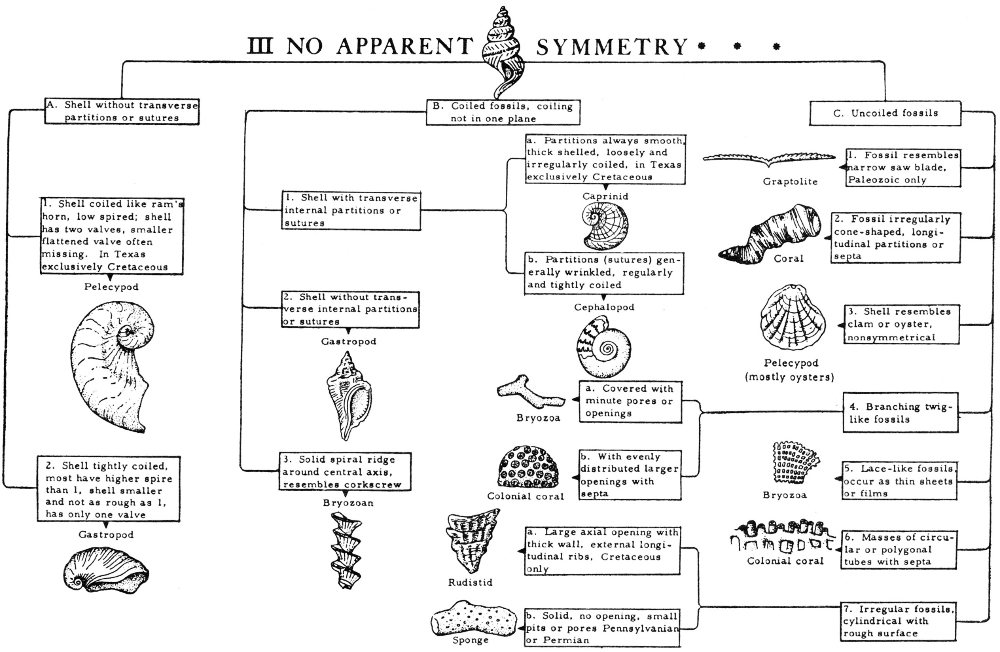
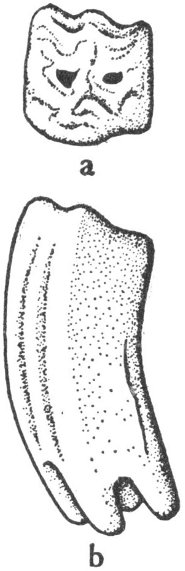
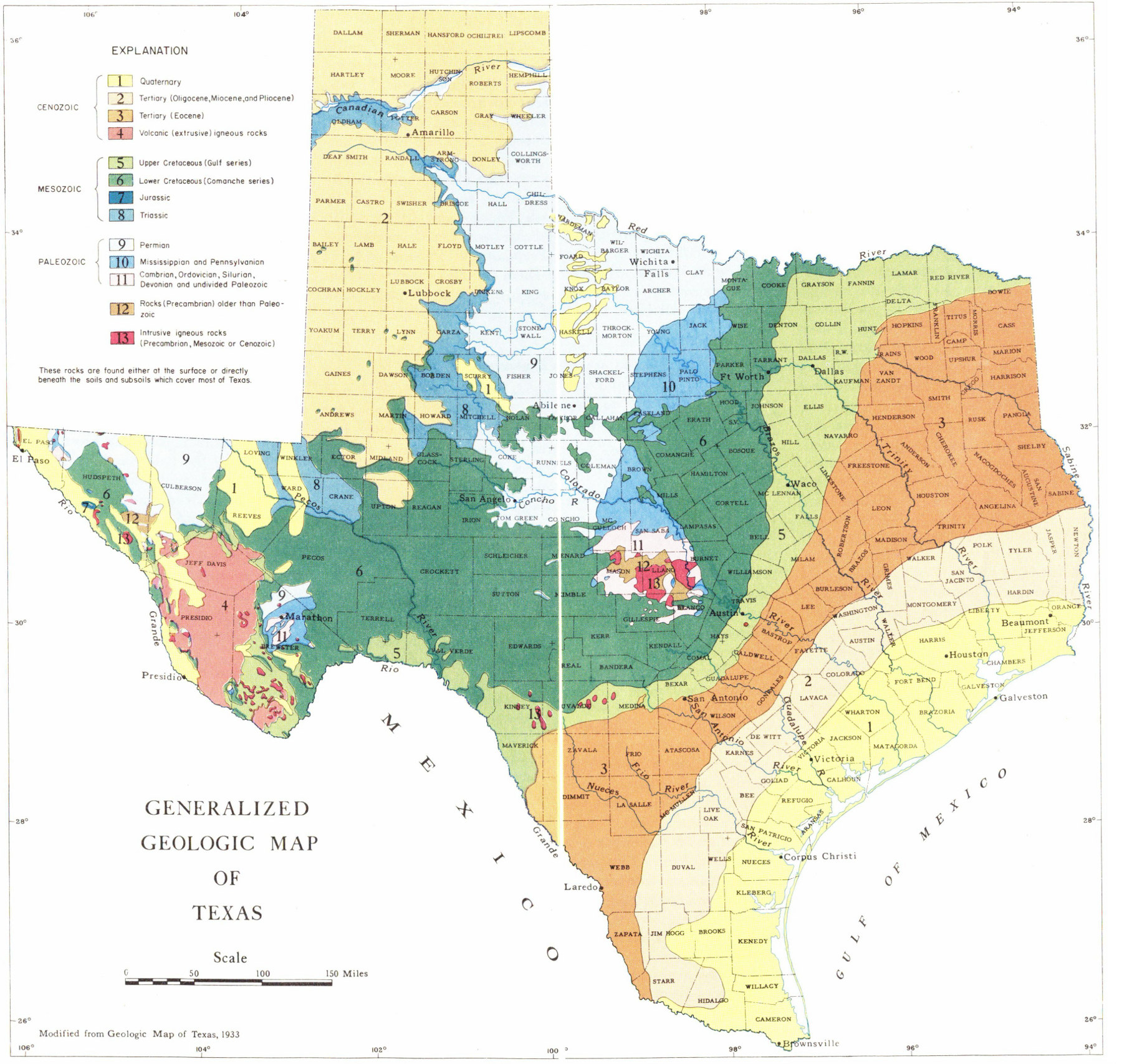{kind=link}